Мир будущего - В. А. Карачунский Философия движения - Мозг как динамическая система. Частные вопросы организации нервной системы.
1. Вступление
Материал, изложенный в данном разделе, должен показать эффективность принципов организации процесса движения в применении к анатомии и физиологии нервной системы человека. Мы будем исходить из общепринятых данных об анатомической схеме мозга и функциональном значении отдельных его частей. В тоже время попробуем показать необходимость и последовательность их появления, как части циклически организованной материи.
С нашей точки зрения, включение эмпирически полученных данных о строении мозга, в концепцию философского плана, позволит их систематизировать. В настоящее время распространено мнение, что философия становиться рудиментом естественных наук, которые перехватывают инициативу ее развития. Это мнение ошибочно. В каждой науке существует логика рассуждений, которая постепенно развивается и объединяет весь собранный материал в едином логическом поле. Философия стоит у истока появления всех наук и то, что она потеряла контроль над их содержимым, превращает эти науки в самостоятельные философские школы, вступающие в противоречие друг с другом, по ряду вопросов. Вопрос должен стоять иначе – необходимо найти логику, общую для всех наук и, тем самым, усовершенствовать саму философию.
Математическая логика способна, на основе того или иного принципа, связать любые данные. В то же время, интерес представляют логические конструкции, имеющие практическое значение, реализуемое в системе координат человеческого сознания, которое представляет собой упорядоченное представление об окружающем мире, то есть философию. Математику без философии можно уподобить компьютеру без оператора. Специалист в узкоспециальном разделе знаний, относящихся к организации и функционированию мозга, не склонен распространять логику своих рассуждений даже на соседние дисциплины, не говоря уже о создании философской системы. Множество “локальных логик” нуждаются в разработке принципов, способных объединить их в единую систему. Например, в нашей работе, принципы организации двигательного процесса, на первый взгляд не связанные с вопросами функционирования мозга, помогают решить ряд проблем, связанных с теоретическим осмыслением вопросов неврологии там, где этого не удается при стихийном обобщении экспериментального материала.
2. Тонус скелетной мускулатуры
Тонус мышц – это сочетание активности отдельных мышечных волокон, мышц и их объединений, фон, на котором происходит тот или иной поведенческий акт. Мышечный тонус отражает уровень активности нейронов моторного мозга и дает зримое представление об узоре информации в этом участке нервной системы. Проекция узора информации из афферентных участков моторного мозга в сенсорный мозг, есть “темное мышечное чувство”, ощущение нашего “Я”. Перестройка тонуса отдельных мышечных волокон (замена одного узора информации на другой), проявляется феноменом поведения. Функциональной единицей моторного отдела нейронной сети является рефлекторное кольцо.

Кольцевое движение импульсов, постоянно присутствующее в нервной системе, является динамической моделью активности мышцы, с которой она связана.
Существование рефлекторного кольца определяется обратной связью, с участием своеобразного рецепторного аппарата - мышечного веретена ( В) и поддерживается определенным тонусом мышечных волокон ( М) , с которыми связан узел моторной сети. Сокращение мышцы совпадает с “выбросом” избытка энергии из моторной нейронной сети и ограничением поступления энергии по обратной связи. У расслабленной мышцы, рецепторный аппарат мышечного веретена находиться в максимально раздраженном состоянии, а у сокращенной мышцы, активность его минимальна. Такое соотношение сложилось в процессе эволюции и сделало возможным кольцевой структуре импульсов, временно превращаться в канал выхода энергии (возникающий в нервной системе избыток энергии, достигнув группы афферентных нейронов ( АН), выталкивает энергию из рефлекторного кольца, заставляя сокращаться мышцу). В момент сокращения мышцы, уровень энергии в рефлекторном кольце падает, а в момент расслабления, восстанавливается, с восстановлением целостности кольца.
Энергия продвижения импульсов через периферические моторные узлы находиться в динамическом соответствии с активностью мышечной ткани и никогда не достигает крайних значений, благодаря противоположной активности моторных нейронов нервного узла и мышечных веретен. Когда мышца значительно расслаблена, активность мышечных веретен частично восстанавливает ее тонус (ввиду того, что минимум энергии в мышце совпадает с максимумом энергии в мышечных веретенах, между ними, при посредстве нервной системы, устанавливается динамическое равновесие, проявляющееся в частичном сокращении мышцы). Таким образом, суммарное кольцевое движение импульсов в моторной сети, скорость элементов которого периодически меняется, является динамической моделью функционального состояния мышечной ткани.
Как видим, участок мозга, непосредственно связанный с мышечной системой, является носителем особой материи, устойчивого кольцевого движения импульсов, которое воспринимается, со стороны, в качестве тонуса мышц, а изнутри, субъективно, как существенная часть нашего “Я”. Изменение внешней обстановки порождает избыток энергии в нервной системе, который расходуется на изменение скорости циркуляции импульсов в моторной сети и проявляется скачкообразной сменой мышечного тонуса (активность организма во внешней среде).

Кольцевые потоки импульсов образуют в нервной системе структуры, устойчивые во времени и пространстве. Благодаря тому, что веретена находятся в толще мышцы, ее возбуждение оказывает на них тормозное действие, что показано жирной стрелкой.
Волны энергии, постоянно поступающие в нервную систему из внешней среды, периодически модулируют активность эфферентных нейронов ( ЭН ), непосредственно связанных с мышечной системой. По сути дела, вся нервная система, ее сенсорная и моторная части, являются самоорганизующимся, постоянно усложняющимся фильтром на пути энергии внешнего воздействия, поступающей через сенсорные анализаторы. Каждый слой нейронной сети, расположенной над двигательными ядрами ствола и спинного мозга, по-своему модулирует уже имеющийся тонус мышечной ткани или скорость продвижения импульсов по кольцам, связанным с теми или иными мышцами.
Движение импульсов в первом слое моторной сети, в упрощенном виде, можно представить следующим образом:

Движение потока импульсов по кольцу, благодаря мышечному веретену, передает инициативу по модуляции активности эфферентных нейронов непосредственно от рецепторного аппарата мышцы, структурам сенсорного мозга.
В достаточно развитой нервной системе, структура периферических моторных узлов выглядит несколько сложнее. Как показано на схеме, в моторной рефлекторной сети формируются несколько кольцевых структур. Кольцо с участием гамма-мотонейронов и кольцо с участием эфферентных нейронов (которое способно поглощать избыток энергии, поступающий из сенсорного мозга). Волна избытка энергии, появившаяся в нервной системе, нарушает равновесие в кольцевой структуре импульсов, за счет усиления активности эфферентных нейронов. Мышца начинает сокращаться. Мышечные веретена блокируют кольцевой поток импульсов, так как в момент сокращения, она тормозит активность мышечного веретена и кольцевая структура моторного узла, превращается в канал выхода энергии. После сокращения мышцы, в результате активности мышечных веретен, она снова переходит в расслабленное состояние. Кольцо импульсов восстанавливается, становиться способным к поглощению очередной волны энергии из сенсорного мозга (восстанавливается способность моторного мозга “сотрудничать” с сенсорным мозгом).
Чем сложнее нервная система, тем сложнее тонус мышц. Различные состояния мышечного тонуса определяются “совместными усилиями” макро и микроструктур нервной системы (оправой, в которую заключены циклические потоки пульсации). Тонус мышц характеризует процедурную память нервной системы, определяющую поведенческую активность организма во внешней среде. Физиология, рассматривая тонкости взаимоотношения образований связанных с мышечным тонусом, не фиксирует внимания на том, что он является проявлением особой материи, представленной кольцевыми потоками импульсов и образующей динамическую модель мышечной системы (можно образно сказать, что при изучении русла реки, мы не имеем представления о воде в нем протекающей). Система циклически организованных потоков импульсов является прослойкой, посредником, которая превращает энергию внешней среды в энергию мышечного движения, чем достигается восстановление равновесия между организмом и окружающей его средой.
Общепринятое описание сенсорных аппаратов скелетно-мышечной системы (48) создает мнение, что информация от проприорецепторов поступает в нервную систему, где она кем-то используется. Кто адресат информации не уточняется, что направляет весь ход теоретической мысли в ложном направлении. С нашей точки зрения, в нервной системе существуют динамические модели внешнего мира и тканей организма. Поскольку оба источника движения объединяться в среде циклических потоков импульсов, они взаимодействуют между собой и образуют совместный вектор движения. В конце концов, организм и среда приходят к временному равновесию, что описывается, как адаптация организма во внешней среде. Как видим, этот процесс самоорганизующийся и не нуждается в постороннем управлении.
3. Локомоция и произвольные движения
Как показано в предыдущей главе, структура, образованная кольцевыми потоками импульсов в нейронной сети, является местом смешения двух потоков энергии, от тканей организма и внешней среды. В результате такого объединения происходит превращение энергии внешнего воздействия в энергию мышечной активности, что приводит к восстановлению равновесия, между средой и организмом. Именно стихийное восстановление равновесия с внешней средой, является фактором отбора и закрепления в структурах памяти тех или иных форм поведения организма.
Тоническое сокращение мышц (сочетание скорости продвижения импульсов в отдельных кольцевых системах) может быть обозначено, как узор информации. Под давлением внешней среды, варианты тонуса мышц и соответствующие ему узоры информации в нервной системе, периодически сменяют друг друга. Таким образом, локомоцию можно записать как циклическую смену определенного набора узоров информации в нейронной сети:
![]()
Во второй части книги приводилась схема развития отдельных узлов и всей нейронной сети, как формирование барьеров противодействия внешней среде. Была показана послойная эволюция сети, отражающая различные уровни организации мышечной активности.

Структурный элемент процесса локомоции – два кольцевых потока импульсов, объединенных в одном узле и система вставочных нейронов, благодаря которой мышцы не могут быть сокращенными или расслабленными одновременно. Энергия внешней среды, попеременно поступает в ту мышцу, которая находится в расслабленном состоянии.
Локомоция – это автоматически запрограммированная деятельность организма в определенных условиях среды. В перспективе развития вида, можно говорить о формировании филогенетической памяти в нервной системе (появлении послойно расположенных макроструктур, закрепляющих рисунок движения между отдельными мышцами и их объединениями). В перспективе научения, речь идет о формировании микроструктуры нервной системы на протяжении жизни индивида, что проявляется, как усложнение аппарата сочетания врожденных форм локомоции. Таким образом, произвольные движения, это стихийная, прижизненная выработка вариантов сочетания врожденных элементов локомоции, а образовавшиеся, на этой основе, автоматические формы поведения, это условные рефлексы.
Пульсация нейронов первого слоя узлов моторной сети поддерживает существование в нервной системе кольцевых структур, которые находятся в динамическом соответствии с тонусом отдельных мышц. В узлах второго слоя, где нет условий для прямого выхода избытка энергии во внешнюю среду, формируется система вставочных нейронов (второй барьер противодействия), которая попарно объединяют группы клеток, связанных с мышцами-антагонистами. Избыток энергии, поступающий в такой узел, находит выход в той его части, которая связанна с расслабленной мышцей (параллельно происходит активизация вставочных нейронов расслабляющих мышцу антагонист). Сократившаяся мышца теряет возможность выводить энергию из связанного с ней участка нервного узла, поэтому дальнейший выход энергии реализуется в той части нервного узла, которая связана с расслабленной мышечной единицей. Таким механизмом попеременного сброса энергии в мышечную систему, стихийно сложившимся в процессе эволюции, обеспечивается непрерывный выход энергии из нервной системы.
В свете излагаемого материала, механизм эпилептического припадка представляется, как наиболее примитивная форма локомоции, имеющая своим единственным результатом вывод избытка энергии из нервной системы. Резкое увеличение энергии в нервной системе сопровождается стиранием узора информации в отдельных нервных узлах, т.к. все нейроны максимально возбуждаются. Исчезает количественная градация в динамической структуре колец потоков импульсов в нейронной сети, которая обеспечивала сложные локомоторные акты. В создавшейся ситуации, энергия находит выход только через группу нейронов, непосредственно имеющих выход в мышечную систему. Процесс этот воспринимается внешним наблюдателем, как тоническое или клоническое сокращение мышц.
Описанные Э. Кэнделом командные нейроны (70) функционируют по принципу, который следует из логики наших рассуждений. Они обеспечивают колебательный выход энергии, через моторную часть нервной системы. В более сложно устроенной нервной системе речь идет не об отдельных командных нейронах, а об их объединениях – нервных узлах. Следует еще раз подчеркнуть, что все процессы в нашей работе рассматриваются в контексте самоорганизующегося процесса, восстановления равновесия между организмом и внешней средой.
Мышечная система, идеальное “изобретение природы”, устройство для отвода избытка энергии из тканей организма, самый универсальный барьер противодействия окружающему миру. Моторный мозг, это самоорганизующаяся система, которая, по мере эволюции, запоминает все более совершенные, стихийно сложившиеся варианты локомоции. Она выводит поступающую в нервную систему избыточную энергию внешней среды, тем самым, охраняя ее структуры от разрушения. Между изменениями внешней среды и поведением организма, локомоцией, существует динамическое равновесие. Поскольку оно постоянно нарушается, а в арсенале памяти не всегда есть уравновешивающие программы поведения, появляется особая форма локомоции, выражающаяся в искажении уже существующих двигательных программ и их необычном сочетании. По сути дела, она есть не что иное, как хаотическое движение блоков, целых программ поведения, а не отдельных мышечных групп (в любом случае, циклическая форма взаимоотношений организма и среды заменяется линейной формой движения). Исходом эволюции (линейного движения) отдельных программ поведения, будет случайная адаптация организма в изменившихся условиях среды, которая, при многократном повторении, изменяет структуру рефлекторной сети (консолидируется) и становиться не случайным явлением, а закономерным.
В процессе эволюции, в моторной сети формируются участки, связанные с мышечными группами, сокращение которых не влияет на равновесие организма и среды. К таким мышечным группам, которые специализируются на хаотических движениях, следует относить мышцы руки (имеется в виду участие руки в жестикуляции, а не в трудовых программах поведения), мышцы голосового аппарата, мимические мышцы. Хаотические движения, являются, по сути дела, самой примитивной и в тоже время наиболее универсальной программой вывода избытка энергии. Со временем, участки моторной сети и связанные с ними мышечные группы, специализируются на “бесполезной форме поведения”. Появление нескольких уровней организации моторной сети способствует запоминанию наиболее часто используемых сочетаний хаотической активности. Хаотические движения становятся более упорядоченными. Появляется определенный их стереотип – исследовательское поведение или ориентировочный рефлекс. Одно из самых организованных проявлений ориентировочного рефлекса представлено мыслительным процессом. Верхние этажи моторной сети, где сосредоточены штаммы исследовательского поведения, выработанные при жизни индивида, представлены фронтальной корой. Более примитивные, врожденные стереотипы ориентировочного поведения, расположены в подкорке. Таким образом, в тех случаях, когда локомоция становиться неэффективной, в плане выведения избытка энергии из нейронной сети, происходит активация хаотической формы двигательной активности, которая, в развитой нервной системе, представлена ориентировочным рефлексом.
Е.Н. Соколов, изучая ориентировочный рефлекс, предположил, что в процессе повторения индифферентного стимула, без специального подкрепления, в мозге формируется его нейрональная модель (120). По нашему мнению, появление модели в нервной системе, это универсальный принцип ее взаимоотношений с внешней средой. Только модель эта не статическая, а динамическая. Она уравновешивает определенные изменения, происходящие во внешней среде. Ориентировочный рефлекс, наряду с хаотической формой поведения, это наиболее универсальный способ вывода энергии из мозга. Модель стимула, в нервной системе (автоматическая программа поведения), не “тормозит” ориентировочный рефлекс, а выводит избыток энергии, являющийся его пусковым механизмом. Соколов связывает модель стимула с нейронами “новизны” и “тождества” гиппокампа (121). Логика наших представлений приводит к выводу, что модель стимула в нервной системе не может существовать вне поля сознания и его свойств, следовательно, в ее формировании принимают участие все нейроны нервной системы. То, что в гиппокакмпе существуют нейроны, реагирующие на любое изменение внешней обстановки, будет объяснено по ходу дальнейшего изложения с несколько других позиций. Предварительно можно сказать, что гиппокамп, участок мозга, принимающий участие во всех изменениях, происходящих в нервной системе.
“Побуждение к движению” принято объяснять активностью подкорковых и корковых мотивационных зон (57). Формально - это верно, но создает путаницу в понимании вопроса. Во-первых, в нервной системе постоянно существует поле образованное циклической формой движения импульсов, которое испытывает давление не только со стороны тканей организма, но и со стороны сенсорных анализаторов. Во-вторых, все движения в тканях организма, прямо или косвенно связаны с динамикой окружающего мира. Мотивация, как будет далее показано, не руководит поведением, а выражает опыт взаимоотношений среды и организма накопленный в процессе эволюции.
4. Мозжечок, как орган, регулирующий скорость выхода избытка энергии
Попробуем рассмотреть мозжечок, как звено в системе, нейтрализующей избыток энергии в мозге. Моторная часть нервной системы связана с тканевыми образованиями, которые способны поглощать энергию. К ним относятся скелетная и гладкая мускулатура, а так же гормональная ткань. Энергия поступает в моторную сеть, через ее афферентные участки, непрерывным потоком. Даже при наличии в моторной сети потенциальной возможности, практическое осуществление выхода избытка энергии сталкивается с определенными трудностями.
Как следует из логики организации и функционирования моторной нейронной сети, энергия выводиться из нее определенными порциями, в момент прерывания кольцевого потока импульсов в ее наружном слое (эфферентные зоны, связанные с сокращающимися мышцами, становятся “воронками” поглощения энергии). Такое положение дел допустимо в моторной сети, имеющей один уровень организации. В многослойной моторной сети, где реализуются сложные программы поведения, имеет значение не только факт продвижения энергии, но скорость, с которой это продвижение происходит (поступление энергии в моторную сеть, происходит не в момент сокращения мышц, а в момент расслабления их антагонистов).
Процесс расслабления мышцы порождает волну избытка энергии (от мышечных веретен). При растяжении изолированной мышцы, поступление энергии, в афферентные участки моторной сети, происходит взрывообразно, что не всегда соответствует возможности ее выхода. Таким образом, избыточная скорость поступления энергии по обратной связи, в процессе изменения тонуса скелетной мускулатуры, вызывает ее “застой” в афферентных участках моторных узлов. Это способствует появлению нейронов, которые отдают породивший их избыток энергии, как тормозной медиатор (избыток энергии находит выход в модуляции активности мотонейронов). Эффективность медиаторов, вызывающих возбуждение эфферентных нейронов, становиться меньшим и процесс сокращения-расслабления мышцы приобретает плавный характер.
Таким образом, формируется структура, нейтрализующая компонент избытка энергии, в афферентных участках моторной сети, возникающий в результате чрезмерно быстрого растяжения мышцы. Со временем, такая структура преобразуется в многослойную сеть нейронов, которая надстраивается над афферентными участками моторной сети и регулирует активность процесса изменения тонуса мышц (фактически, эта структура становиться барьером противодействия на пути энергии, поступающей по обратной связи от скелетной мускулатуры).
Как стало ясным из хода изложения, структурой, нейтрализующей избыток энергии, поступающего в моторную сеть при расслаблении мышц, является мозжечок. Важно отметить, что мозжечок формируется, как самоорганизующаяся система, уравновешивающая избыток энергии в моторной сети. Мозжечковые воздействия делают плавным процесс поступления избытка энергии обратной связи в моторную сеть, за счет торможения активных участков моторной сети (не дают быстро расслабляться мышцам антагонистам). По сути дела, формирование мозжечка происходит по тем же законам, что и появление тормозных элементов, в любом сенсорном анализаторе. Вставочные нейроны ограничивают поступление энергии, через рецепторный аппарат анализатора. Только в случае с мозжечком, мы имеем дело не с наружной внешней средой, а с внутренней (в качестве рецепторного поля выступают мышечные веретена) и ограничивается не процесс поступления энергии, а его выход во внешнюю среду.
Сеть тормозных нейронов в моторной сети, специализирующихся на энергии обратной связи, является структурой филогенетической памяти, в которой, процессе воспроизведения, формируется негативная копия различных состояний тонуса организма. Любая форма мышечной активности модулируется тормозными воздействиями мозжечка по принципу: “сила действия соответствует силе противодействия”.
Элементарная схема взаимоотношений структур моторной сети, нейтрализующих избыток энергии, поступающий по обратной связи, выглядит следующим образом.

Избыток энергии в моторной сети, при избыточно быстром расслаблении мышцы, формирует скопление вставочных нейронов, которые вырабатывают тормозной медиатор, который снижает активность расслабления мышцы.
Сокращение определенного количества мышц и соответствующее расслабление их антагонистов, порождает вону избытка энергии в моторной сети. В результате послойного развития тормозной сети, которая уравновешивает этот избыток энергии, формируется мозжечок. Афферентная информация от мышечной и гормональной тканей организма, запускает тормозные воздействия мозжечка. Они модулируют эффективность возбуждающих медиаторов в участках моторной сети, связанных со структурами находящимися в состоянии расслабления.

В основе поведения лежит быстрая смена тонуса мышечной системы. Процесс уравновешивается активностью мозжечка, которая делает его плавным и позволяет формироваться сложным программам поведения, с участием многослойной структуры нейронной сети.
Принципиальную схему мозжечка, как части нервной системы, можно представить следующим образом.

Мозжечок получает энергию от входов в узлы моторной сети, связанные с активностью той или иной программы поведения и отдает ее, в виде тормозного медиатора, в узлы связанные с мышцами антагонистами, которые участвуют в текущей программе поведения.
На схеме показана моторная нейронная сеть, связанная со скелетной мускулатурой. Во всех ее узлах появляется один и тот же узор информации, уравновешивающий данную внешнюю обстановку, например гомонкулюс с поднятой рукой. Если гомонкулюс опустит руку, то во всех узлах моторной сети произойдет то же самое. Предположим, на входе в структуры мозжечка появиться узор информации, гомонкулюс с поднятой рукой. На выходе из мозжечка, в моторную сеть возвращается тот же самый узор информации, что и на входе в него, только действие его будет не возбуждающим, а тормозящим (поскольку нейроны, на которые накладывается узор информации от мозжечка, уже находятся в состоянии возбуждения, знак медиатора, который он вырабатывает, может быть только тормозным). Действие мозжечка можно сравнить с вязкой средой, которая не дает организму совершать резкие движения. Этот процесс выражен тем сильнее, чем резче происходит сокращение мышечной ткани.
В процессе эволюции мозжечок становится местом, где собираются и консолидируются все узоры информации, которые уравновешивают любой вариант тонической перестройки мышечной системы организма (каждый, вновь появившийся вариант равновесия со средой, представлен определенным узором информации). Поскольку мозжечок является многослойной структурой, ряд слоев которой имеет выход только в собственные внутренние структуры, у нейронов верхнего слоя мозжечковой нейронной сети развит нейропиль, третий барьер противодействия внешней среде. Особо показательны в этом плане клетки Пуркинье, дендриты которых напоминают крону дерева. Гипертрофированная мембрана данных нейронов позволяет уравновесить тончайшие нюансы мышечной активности организма. Таким образом:
мозжечок – это образование моторной сети, уравновешивающее избыток энергии, возникающей при появлении обратной афферентации от тканей организма

Графики характеризуют скорость выхода энергии из изолированной мышцы и скорость выхода энергии из мышцы, получающей модуляцию со стороны мозжечка.
5. Вестибулярный аппарат
Все живое на земле существует, как часть общей динамической системы (субъективно мир состоит из отдельных объектов, отношения между которыми рассматривается в контексте взаимодействия). Изменения среды вызывают ответную реакцию в тканях организма и наоборот, протекающие в организме процессы оказывают влияние на среду вокруг организма. При перемещении организма в пространстве в нем возникает избыток энергии, связанный с действием гравитационных сил. Возникнув вблизи от элементов мышечной ткани, такой избыток энергии находит выход в хаотическом движении. По мере эволюции, участок входа энергии преобразуется в сенсорный орган (вестибулярный аппарат), а мышечная ткань приобретает изначальную дифференциацию. Следует отметить, что рецепторное поле, регистрирующее данную модальность, расположено в тканях и не нуждается прямом контакте с внешней средой.
Таким образом, появление рецепторного поля, поглощающего энергию гравитационного воздействия, лежит у истоков дифференциации мышечной ткани, активность которой презентовала первую программу поведения организма, обеспечившую стабильное положение организма в пространстве (со временем, мышечная система стала не только противостоять силам гравитации, возникающим при перемещении организма, но и учитывать скорость этого перемещения). По мере эволюции участка нейронной сети, связанного с вестибулярным аппаратом, активность мышечной системы становилась все более упорядоченной.
Вестибулярный аппарат, является сенсорным анализатором, способным регистрировать такие свойства окружающей среды, как гравитация, что субъективно проявляется чувством движения вперед, назад, влево, вправо, вверх, вниз, в их различных сочетаниях. Неочевидность восприятия гравитации, в качестве сенсорной модальности, обусловлено тем, что система противодействия в высокой степени автоматизирована и не нуждается в дополнительной коррекции со стороны коры мозга. Избыток энергии, поступающий в моторную сеть от вестибулярного аппарата и тканей организма, в основной своей массе не выходит за ее пределы. Поэтому он, будучи нейтрализован внутренними механизмами моторной сети, поступает в сознание, как трудно дифференцируемое ощущение нашего “Я”. Представления о положении в пространстве присутствует в сознании, как нечто само собой разумеющееся, фон на котором появляются более поздние сенсорные модальности (для его осознания необходимо развитие абстрактного мышления).
Вестибулярный аппарат, один из древнейших сенсорных анализаторов. Он имеет представительство на всех этажах нервной системы. Уже на уровне ствола мозга вестибулярный аппарат тесно контактирует с другими сенсорными анализаторами, так как эффективность их работы находиться в прямой зависимости от их стабильного положения и перемещения организма в пространстве. По ходу изложения материала, главным приоритетом является соблюдение принципа, согласно которому все участки мозга рассматриваются, как самоорганизующуюся система (процесс становления равновесия между организмом и окружающей средой).
С этих позиций схема вестибулярной системы представляется следующим образом. При изменении положения организма в пространстве на него начинают действовать гравитационные силы, в результате чего меняется уровень пульсации нейронов рецепторного поля. От рецепторов избыток энергии поступает во второй слой нервных узлов и далее находит выход в мышцах, сокращение которых изменят тонус мускулатуры определенных мышечных групп, позволяя организму сохранить исходное положение в пространстве. Вестибулярный аппарат, по логике своего развития и функционирования, напоминает мозжечок, вырабатывающий возбуждающий медиатор.

Энергия внешней среды, поступая в вестибулярный аппарат, сбрасывается в группу мышц, активация которой нивелирует гравитационное воздействие.
Таким образом, приходим к выводу:
вестибулярный аппарат, это сенсорный анализатор, реагирующий на изменение сил гравитации, действующих на организм и модулирующий активность мышечной системы, в плане противодействия изменениям положения организма в пространстве
6. Вегетативная нервная система
Факт вегетативной афферентации тканей долгое время не признавался многими исследователями. Постепенно был собран достаточный материал свидетельствующий о том, что в вегетативной нервной системе присутствуют как сенсорные, так и моторный звенья. Так А.Д. Ноздрачев пишет (123):
“Автономная и соматическая рефлекторные дуги построены по одному и тому же плану и состоят из чувствительного, ассоциативного и эфферентного звеньев… Таким образом, автономная нервная система по составу нейронных звеньев дуги, нисколько не отличается от соматической, совершенно не характеризуется “чисто эфферентной природой”, как это ей приписывалось до недавнего времени.”
С точки зрения наших представлений о нервной системе, данный факт не подлежит сомнению. Вегетативная нервная система является частью моторной нейронной сети, имеющей многослойное строение, которая уравновешивает внутреннюю внешнюю среду. По ходу эволюции, каждый слой этой системы (интрамуральные элементы, сплетения, вегетативные узлы и так далее), был высшим этажом регуляции процессов, происходивших в тканях организма и не мог обходиться без обратной связи. Внутренняя внешняя среда, которой противодействует вегетативная часть моторной сети, расположена непосредственно вокруг клеток организма (кровь, лимфа, тканевая жидкость). В ее динамике отражаются все процессы, происходящие в организме. Возникающий при этом избыток энергии улавливается разнообразными рецепторами и возвращается обратно, в виде активности гладкой мускулатуры сосудов, внутренних органов, железистого аппарата экзоэндокринных и эндокринных желез.
Среда, с которой имеет дело вегетативная система, в отличие от среды окружающей организм, имеет конечные размеры и относительно небольшой набор функциональных состояний. Появляющийся в ней избыток энергии хорошо уравновешивается сравнительно несложной структурой вегетативной сети нейронов и не нуждается в значительном развитии афферентных элементов. Соответственно, верхние этажи вегетативной системы являются малодифференцированными и в сознании представлены неясными, трудно характеризуемыми ощущениями. В этом плане, вегетативная нервная система напоминает нижние этажи моторную нейронной сети, которые уравновешивает контактирующую с ней внешнюю среду в автоматическом режиме и не нуждаются в чрезмерном развитии своей афферентной части. Принципиальная схема эволюции парасимпатической нервной системы, может быть представлена следующим образом.

В структуре парасимпатической нервной системы отражаются различные уровни движения, происходящие в тканях организма
Первый слой нервных узлов можно сравнить с эндокринными железами, которые обладают локальным действием и обеспечивают определенный диапазон активности связанных с ними участков ткани (гладкомышечных элементов и желез). Последующие слои обеспечивают нарастание уровня коммутативной активности органов и их объединений (для второго слоя нейронной сети параметры первого являются внешней средой, поэтому он “учиться” противодействовать движению которое отражает коммутативную активность групповых объединений узлов первого слоя). Между участком внутренней среды организма и рядом расположенным нервным узлом, между узлами первого уровня и узлами следующих слоев, образуются кольца движения импульсов, скорость перемещения которых отражает уровень функциональной активности участка ткани, органа, системы органов. Парасимпатический участок моторной сети обеспечивает общий вектор поведения клеток, органов и систем органов, противодействующий изменению параметров среды вокруг клеток организма.
Активное противодействие организма окружающей среде, за счет развития скелетной мускулатуры и усложнения поведения, модулирует состояние внутренней среды организма и предъявляет повышенные требования к функционированию парасимпатической системы, а также тканям с ней связанным. Это приводит к появлению избытка энергии в верхних ее этажах. Последний находит выход, в появлении и развитии нейронной сети, которая осуществляет модуляцию активности парасимпатической системы на входе (перестройка активности тканей способствует более эффективной работе скелетной мускулатуры, в соответствии с изменившимися условиями среды). Вновь образовавшаяся сеть нейронов образует симпатическую нервную систему, своеобразный “пирамидный путь”, управляющий внутренней средой организма. Схематически процесс взаимодействия двух частей нервной вегетативной системы, можно представить следующим образом.

Симпатическая нервная система развивается из афферентных зон верхнего этажа парасимпатической нервной системы. Эфферентная ее часть, принимая избыток энергии от соматической части нервной системы, находит ему выход в “стрессовой перестройке” тканей организма, уравновешивая воздействия окружающей среды
Вегетативная нервная является структурой филогенетической памяти, которая преобразует активность единичных клеток в системное поведение клеток всего организма. Перестройка активности тканей, под воздействием симпатической нервной системы, способствует более эффективной работе скелетной мускулатуры, в плане более высокого уровня равновесия между внешней средой и организмом. Таким образом:
Внутренняя среда организма способствует появлению в тканях организма автономной нервной системы, которая модулирует процессы, протекающие во внутренней среде организма
Парасимпатическая нервная система обеспечивает программу поведения клеток, которая поддерживает параметры внутренней среды организма
Симпатическая нервная система обеспечивает программу поведения клеток ориентированную на противодействие факторам внешней среды, не имеющими прямого контакта с клетками организма
7. Гипоталамус
Вокруг нервной системы существует две внешние среды, с которыми она контактирует через рецепторные аппараты. Первая представлена средой вокруг организма, вторая средой внутри организма (во второй части книги было предложено назвать их наружной и внутренней внешней средой). Важно подчеркнуть, что кровь и тканевая жидкость, среда общая для всех клеток организма, имеет ограниченные размеры и контактирует с наружной внешней средой, через ткани организма. Поскольку ДНК определяет логику процессов, протекающих в клетках данного организма, которые контактируют с его внутренней средой, последней будет свойственна логика движения ДНК. Динамические особенности крови и тканевой жидкости передаются спинномозговой жидкости, которая содержится в желудочках мозга и отделена от кровяного русла своеобразным фильтром, гематоэнцефалическим барьером (можно сказать, что логика динамики ДНК отражается в ней в упрощенном, абстрагированном виде).
Гипоталамус, отдел моторного мозга, имеющий сенсорные входы от спинномозговой жидкости, содержащейся в третьем желудочке и гипофиза, нейросекреторного образования, отражающего законы гомеостаза. Гипофиз и основные железами внутренней секреции образуют динамическую систему, сформированную по типу обратной связи, которая противодействует изменениям параметров внутренней среды организма, модулируя функциональное состояние отдельных тканей гуморальными воздействиями. Из сказанного можно сделать вывод, что гипоталамус представляет собой участок мозга, отражающий логику движения ДНК (внутренней среды организма). Таким образом, циклические потоки импульсов, существующие в гипофизе и гипоталамусе, являются динамической моделью процессов, протекающих во внутренней среде организма, то есть представляют копию ДНК. Гипоталамус, в свою очередь, испытывает на себе влияние сенсорного мозга, циклические потоки импульсов которого отражают динамику внешней среды. Следовательно, модуляция функционального состояния гипоталамуса воздействиями внешней среды, передается через кровь и спинномозговую жидкость, на ткани организма, вызывая их функциональную и структурную перестройку. Поскольку внутренняя среда организма контактирует с половыми клетками, имеющими половинный набор хромосом, изменение ее параметров создает возможность для закрепления на генетическом уровне параметров тех участков организма, где под воздействием внешней среды произошла функциональная и структурная перестройка. Как видим, существует вполне “легальный” путь плавной адаптации генетического материала к условиям внешней среды, а о мутациях следует думать только в контексте партеногенеза. С данной точки зрения, учение Ж.Б. Ламарка во многом справедливо (127).

Между двумя динамическими системами, ДНК и внешней средой, образуется целая система слоев-посредников. В процессе становления между ними равновесия происходит плавное изменение генетического материала.
Гипоталамус представляет участок моторной нейронной сети, в каждом слое которой отражается определенный уровень движения, существующий в кровяной ткани. Первый слой (тканевые элементы вегетативной нервной системы) уравновешивает наиболее динамичные параметры внутренней среды организма, второй слой, принимая избыток энергии от первого слоя, уравновешивает динамические соотношения, складывающиеся между групповыми объединениями элементарных параметров этой среды и так далее. Принципиальная структура гипоталамуса выглядит следующим образом.

В гипоталамусе и гипофизе запоминаются параметры, которые отражают динамику внутренней среды организма. В ответ на афферентную информацию, структура гипоталамуса уравновешивает определенный уровень движения внутренней среды, согласуя свои действия с процессами сенсорного мозга. Афферентная информация от спинномозговой жидкости усиливает адаптационную роль гипоталамуса по параметрам, существенно нарушающим гомеостаз.
Гипоталамус, это своеобразная моторная сеть. Условно можно сказать, что первый слой его нервных узлов является моделью наиболее динамичных параметров внутренней среды организма, длительность существования которых изменяются минутами, часами. Например, изменения гомеостаза во время приема пищи, физической нагрузки и так далее. Второй слой, уравновешивает более медленные процессы, с интервалом колебания в пределах 24 часов (сон, бодрствование). Третий слой уравновешивает колебания параметров внутренней среды с интервалом месяц и более, гормональные циклы, сезонные колебания параметров внутренней среды организма. Четвертый слой уравновешивает еще более медленные биоритмы, рост, половое созревание, процесс старения и так далее (деление на слои проводиться в рамках рассматриваемой теоретической модели и нуждается в экспериментальном подтверждении).
Вышеизложенный материал позволяет представить формирование в многоклеточном организме систем управления, как процесс становления равновесия между организмом и внешней средой. Первые, гуморальные системы управления появились тогда, когда в групповом объединении клеток появилось замкнутая полость. Общее пространство всех клеток организма, внутренняя внешняя среда, ввиду ограниченных размеров, контактируя с гуморальными факторами, продуцируемыми отдельными клетками, была способна менять свои параметры. В тех местах, где динамика изменений внутренней среды была наиболее высока, наряду с функциональными изменениями, происходили структурный перестройки, то есть начиналась дифференцировка тканей. Таким образом, в процессе становления равновесия между внутренней средой организма и его клетками, появились скопления клеток (секреторные железы, вырабатывающие гормоны), уравновешивающие активность отдельных тканевых образований. Объединение инкреторных желез в единую систему, знаменовалось появлением гипофиза, центра гуморального управления организмом, который уравновесил динамику внутренней среды организма. Развитие парасимпатической нервной системы, преобразовало управление клеточными образованиями в процесс более точечный и целенаправленный. Третий этап эволюции систем управления связан с формированием системы модуляции параметров внутренней среды организма, с учетом внешних воздействий (симпатическая нервная система). Дальнейшая эволюция нервной системы ориентирована на противодействие удаленным элементам окружающей среды и знаменуется развитием моторного и сенсорного мозга, управляющего скелетной мускулатурой.
Моторная рефлекторная сеть является областью мозга, которая уравновешивает те или иные факторы среды в автоматическом режиме. Избыток энергии в нее поступающий, без проблем находит выход из нервной системы, поэтому афферентная ее часть, в сравнении с сенсорным мозгом, сравнительно мало дифференцирована.
8. Миндалина
Несмотря на обширность материала, собранного в современной физиологии и разнообразие теоретических положений, нередко возникают трудности с осмыслением элементарных понятий, что объясняется отсутствием достаточной философской концепции, дающей возможность систематизировать существующую информацию. Очень близко к ее созданию подошел А.А. Ухтомский. Его теоретические исследования сущности рефлекса, идеи связанные с доминантой, становятся яснее в парадигме нашего подхода. Так А.А. Ухтомский пишет (18):
“Рефлексы – это следы прежней деятельности и прежних выработок, какие были присущи организму в его приспособлении к среде”
Следует понимать, что рефлексы это зафиксированные в памяти способы противодействия среде (состояние динамического равновесия).
“… рефлекс как таковой, то есть остаток от прежней целостной деятельности организма, не может содержать в себе ничего нового, “творческого”! Он лишь инструмент в руках творческого прибора – организма в целом”
В данном высказывании содержится нерасшифрованная идея о рефлексе, как составляющей общего поля сознания, динамические процессы в котором, при посредстве окружающей среды, порождают новые рефлексы.
“Рефлекс – это, скорее, порядок, в котором совершается акт, а не характер его по содержанию”
“… целое не результат складывания рефлексов, но интеграция моментов действия”
То есть рефлекс это субъективное, одностороннее восприятие взаимоотношений организма со средой, сутью которого является взаимодействие двух систем, внешней среды и поля сознания.
Анализируя теоретические проблемы нейрофизиологии, Батуев А.С. предлагает четыре основных принципа работы мозга – принцип рефлекса, принцип доминанты, принцип отражения, принцип системности (18). Используемый в данной работе подход, к анализу процесса движения, позволяет все принципы рассматривать как производные циклической формы движения. Показателен, в этом плане, участок моторной сети, называемой миндалиной. Миндалина это область, где консолидируются программы противодействия среде, составленные из отдельных блоков локомоторных актов, к которым относятся пищевое, оборонительное и прочие виды врожденных форм поведения.
Идею отражения наглядней рассматривать, как процесс взаимодействия динамических систем (организм – среда), стремящихся к равновесию. Системность вытекает из того, что две движущиеся системы, поневоле образуют одну совместную. По ходу существования организма, в нем накапливается иерархия циклических форм движения, уравновешивающих все большее количество внешних факторов. Одним их этапов равновесия становиться появление нервной системы и формирование в ней миндалины, активность которой уравновешивает определенный уровень взаимодействия со средой. Миндалина формируется методом проб и ошибок, очень медленно, в процессе смены поколений. (Можно говорить о рефлексе, который вырабатывается в течение нескольких столетий). Поскольку процесс эволюции миндалины не может быть охвачен нашим сознанием в целом, во времени и деталях, мы воспринимаем его в парадигме поведения, фрагментарно, как отдельные рефлексы, связь между которыми не всегда очевидна. Тот же процесс, но воспринимаемый, как регистрируемые участки активности в нервной системе, дает субъективное представление о доминанте. Доминанта, это перераспределение активности нейронов и их объединений, при вновь образовавшемся равновесии организма со средой, с точки зрения возбудимости. Для полноты картины, необходимо учитывать участки нейронной сети, где активность снижается.

Миндалина часть организма, появляющаяся в нем на определенном этапе становления равновесия, между организмом и средой, конгломерат автоматической (процедурной) памяти.
Таким образом, миндалина, это участок моторной сети, представленный многослойной узловой структурой, где сосредоточена филогенетическая память, обеспечивающий сложные инстинктивные формы поведения: пищевое, оборонительное и так далее. Волны избытка энергии, поступающие в различные участки миндалины, изменяют ее структуру и приводят к формированию соответствующих поведенческих форм активности, восстанавливающих нарушенное равновесие организма и среды.
9. Вегетативная система сознания
В 1949 году Дж. Моруцци и Х. Мэгун, занимаясь проблемой сна, открыли в стволе мозга неспецифическую систему, мезэнцефалическую ретикулярную формацию, оказывающую активирующее влияние на кору мозга. Дальнейшие исследования привели к открытию других активирующих систем – неспецифической таламической системы и активирующих структур заднего гипоталамуса. В дальнейшем, в целом ряде работ было показано, что отдельные участки ретикулярной формации оказывают тормозящее влияние. Учитывая полученные данные, Дж. Росси и А. Цанкетти предположили существование в стволе мозга двух антагонистических механизмов, десинхронизирующего и синхронизирующего. Данная работа ставит перед собой задачу представить процесс появления активирующих структур мозга, с точки зрения самоорганизующейся системы, стремящейся к равновесию с внешней средой.
Любая ткань организма имеет вегетативную иннервацию, которая обеспечивает прирост или снижение в ней уровня медиаторов, регулируя ее взаимоотношения с внутренней средой организма. Вещество мозга, в этом плане, не исключение. Оно является тканью, зависимой от состояния внутренней среды организма и получает соответствующую регуляцию, со стороны вегетативной нервной системы.
Ткань мозга является видимым материальным образованием. Однако в мозге, помимо нее, присутствует невидимая материальная структура, поле сознания, образованное циклическими потоками импульсов в нейронной сети. Поле сознания, как ткань организма, нуждается в энергетическом обеспечении своих функциональных состояний. Эту роль берет на себя неспецифическая система мозга, своеобразный аналог периферической вегетативной системы. Активность поля сознания определяется скоростью продвижения импульсов по отдельным его участкам. В отличие от прочих тканей организма, активность сознания определяется не только функциональным состоянием нейронов и внутренней среды организма, но и состоянием синаптических контактов. Синапсы регулируют скорость прохождения избытка энергии по нейронной сети (один и тот же узор информации, в зависимости от уровня присутствия медиаторов в нервном узле, может проходить через него с различной скоростью).
Таким образом, скорость прохождения потоков информации, по структурам нервной системы, регулируется неспецифической системой мозга. Ее появление происходит в местах с избыточной функциональной нагрузкой, в виде дополнительной сети вставочных нейронов, которые захватывают избыток энергии в узле, возвращая его обратно, как возбуждающий медиатор (избыток энергии активирует вставочные нейроны ретикулярной формации, которые оказывают влияние на нервный узел, увеличивая его пропускную способность). Сенситизация, процесс облегчения синаптической проводимости, является своеобразным вариантом второго барьера противодействия внешней среде, где не тормозной, а возбуждающий медиатор создает условия для уменьшения избытка энергии в нервном узле.

Избыток энергии в нервном узле, не имеющем прямого выхода во внешнюю среду, активирует вставочные нейроны, увеличивающие его пропускную способность.
Как следует из логики развития нейронной сети, избыток энергии, образующийся в нервных узлах, формирует из вставочных нейронов ретикулярной формации сеть, которая повышают пропускную способность определенных зон мозга, соответственно текущей ситуации.

Неспецифическая система ствола и таламическая неспецифическая система, облегчают прохождение потока энергии через ствол и подкорковый отделы мозга. Ядра шва, черная субстанция и голубое пятно выборочно усиливают активность центров связанных с определенными программами активности поля сознания. Стриопаллидарная система, действуя по принципу мозжечка, выделяет ГАМК, тормозной медиатор, делающий процесс усиления активности медиаторов плавным.
Попробуем представить принципиальную схему внутренней “вегетативной” системы мозга, управляющей интенсивностью потоков импульсов в нейронной сети. Внутри нервных узлов, или рядом с ними, образуются скопления вставочных нейронов, увеличивающих пропускную способность последних. Эти образования являются первым слоем моторной рефлекторной сети, описанной как ретикулярная формация и управляющей интенсивностью потоков энергии в мозге. По сути дела, ретикулярная формация, аналог филогенетической памяти, в которой записана количественная сторона интенсивности потоков импульсов в нервных узлах. Электроэнцефалограмма регистрирует не изменение узора информации в нервных узлах, а рост интенсивности прохождения в них энергии, поэтому судить на ее основании о содержании сознания, можно лишь косвенно (различное сочетание активности отдельных нейронов узла, с точки зрения электроэнцефалограммы, неотличимы, если интенсивность потоков импульсов одинакова). Второй слой сети развивается на базе первого и отражает более медленные уровни функциональной активности отдельных образований в нервной системе. Узлы этого слоя – ядра шва, черная субстанция и прочие, являются аналогами сплетений вегетативной нервной системы. Они “обслуживают” не отдельные узлы, а их сочетания в коммуникативной программе действий. С этой точки зрения, голубое пятно, активирующее кору мозга, это аналог симпатической части вегетативной системы, модулирующей активность поля сознания, согласно изменившимся условиям среды.
Прирост активности нервных узлов не может гарантировано решить проблему избытка энергии в мозге, поскольку внешняя ситуация изменчива и часто не соответствует поведенческим программам. По этой причине, часть элементов неспецифической системы перестраивается на выработку тормозного медиатора, который, по принципу обратной связи, снижает эффективность воздействия неспецифической системы на основные проводящие пути, уменьшая объем потоков энергии в нервной системе. Таким образом, в неспецифической сети появляются скопления вставочных нейронов, оказывающих тормозное действие.
С появлением коры, масса нервной системы кратно возрастает. Появляются проблемы связанные не только с объемом прохождения энергии, но и со скоростью ее прироста (ускорения). В ретикулярной системе формируется стриатум, образование противодействующее скорости нарастания потоков импульсов в нервной системе, своеобразный “мозжечок” для всех процессов в неспецифической системе мозга. В.Н. Бархатова следующим образом характеризует функцию стриатума (17):
“Важно подчеркнуть, что все афферентные системы стриопаллидарного комплекса оканчиваются только в стриатум, который получает импульсы из 4 основных источников: большинства областей коры мозга, центральных интраталамических ядер зрительного бугра, компактной зоны черной субстанции, ядер шва среднего мозга”
“От стриатум пути идут только к бледному шару и ретикулярной зоне черной субстанции, от которых в свою очередь начинаются основные эфферентные системы стриопаллидарного комплекса”
“Через внутренний сегмент бледного шара и ретикулярную зону черной субстанции осуществляется обратная связь между стриатум, зрительным бугром и корой мозга”
“Важные эфферентные системы внутреннего сегмента бледного шара идут к ядрам ретикулярной формации среднего мозга”
“От наружного сегмента бледного шара массивные проекции идут к субталамическому ядру, поражение которого приводит к развитию грубого гиперкинетического синдрома - гемибализма”
Таким образом, сенсорная энергия от всей “вегетативной системы мозга” проецируется, с учетом топографии, в стриатум и через бледный шар направляется обратно, в виде тормозного медиатора (гамааминомаслянная кислота), который снижает скорость модулирующего воздействия неспецифической системы.
Появление в нервной системе функциональных объединений узлов, обслуживаемых определенным участком неспецифической системы, требует медиаторов вызывающих усиление проводимости именно этого объединения и не затрагивающего активность соседних участков. По этой причине медиаторы в мозге представлены большим разнообразием вариантов. Многие из них, обслуживая разные функциональные системы, становятся в реципрокные отношения (например, дофамин, норадреналин, глутамин с одной стороны и ацетилхолин, серотонин с другой). В итоге получаем следующие выводы:
В нервной системе, по мере ее развития появляется моторная рефлекторная сеть "запоминающая" активность различных участков нейронной сети, "количественная филогенетическая память"
В отличие от филогенетической памяти качественной, обеспечивающей запоминание узора информации (чему способствует развитие отростков нейронов, рост нейропиля), память в моделирующей системе мозга представлена гипертрофией массы "нейронных эндокринных желез", имеющих стереотипное строение
Избыток энергии в нервной системе способствует усложнению неспецифической рефлекторной сети, превращению ее в многослойную структуру, в которой появляются тормозные элементы, регулирующие уровень ее возбудимости
Избыточная скорость поступления энергии в нервную систему, связанная с активностью неспецифической системы мозга, порождает ее избыток, который уравновешивается своеобразным мозжечком, стриопаллидарной системой.
10. Феномен боли в нервной системе
. Характеризуя болевую рецепцию, И.А. Шевелев пишет о том, что до сих пор неясно, есть ли специфические болевые рецепторы (138). Далее им сформулированы две альтернативные гипотезы организации болевого восприятия. Согласно одной из них существуют специфические болевые рецепторы, свободные нервные окончания с высоким порогом реакции. Вторая гипотеза утверждает, что специфических болевых рецепторов не существует, и боль возникает при сверхсильном раздражении любых рецепторов.
С точки зрения излагаемого материала, предпочтительней вторая гипотеза. В узлах нейронной сети существует определенный узор информации, сочетание уровня пульсации отдельных нейронов. Если все нейроны узла максимально возбуждены, в результате переизбытка в нем энергии, узор информации исчезает. Следует обратить внимание, что любые узлы нервной системы, относящиеся к любой модальности, при максимальном возбуждении идентичны в информационном плане (если все точки, составляющие изображение на телеэкране, приобретут максимальную яркость, то получим монотонную светящуюся поверхность). Поэтому ощущение боли обусловлено не особенностями рецепции, а особенностями “информационной картинки”, которая универсальна для любого источника и отражает количественную сторону сенсорных процессов.
Поступление информации в нервную систему, это продвижение энергии по каналам сенсорного анализатора, которое сопровождается формированием барьеров противодействия этому процессу. На входе в анализатор ограничительную роль выполняют вставочные нейроны, вырабатывающие тормозной медиатор (латеральное торможение). На последующих уровнях организации анализатора вставочные нейроны перестраиваются и начинают вырабатывать медиатор, облегчающий продвижение энергии (вещество Р). Если энергии в узле появляется слишком много, активируется тормозная система – вставочные нейроны вырабатывающие энкефалин. Например, по нашему мнению, схему болевой рецепции, предложенную Л. Иверсон (50), можно представить следующим образом:
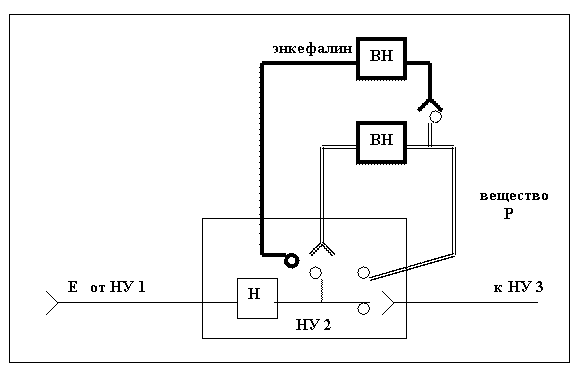
Поток энергии поступает по цепочке нервных узлов. При его избытке в НУ2, активируется первый вставочный нейрон, вырабатывающий вещество Р, увеличивающее пропускную способность узла. Если процесс сенситизации становиться неэффективным, происходит активация тормозных вставочных нейронов, связанных с возбуждающими вставочными нейронами, которые вырабатывают медиатор, снижающий проходимость синапсов.
11. Эмоции, как форма сознания
Один из ведущих специалистов по изучению эмоций, В.П. Симонов, следующим образом характеризует состояние проблемы (113):
“Несмотря на то, что каждый из нас знает, что такое эмоция, дать этому состоянию точное научное определение невозможно. В настоящее время не существует единой общепризнанной теории эмоций, а также точных данных о том, в каких центрах и каким образом эти эмоции возникают и каков их нервный субстрат”
Далее В.П. Симонов приводит категорическое высказывание Б. Райма о том, что современное состояние учения об эмоциях представляет собой:
“…разрозненные знания, непригодные для решения конкретных проблем”
Складывается парадоксальная ситуация, наличие многочисленных гипотез, громадного экспериментального материала, наряду со смутным пониманием того, с чем мы имеем дело. По нашему мнению, трудности в определении сути эмоций, порождает традиционный подход к их рассмотрению, следствие дедуктивного метода, преобладающего в современной науке. Гипотезы, объясняющие, что такое эмоции, находятся в “подчинении” у экспериментальных данных, которые, в виду их чрезвычайно узкой специализации, не могут влиять на кардинальные теоретические подходы, уже используемые для решения поставленных задач. Развитие теоретического подхода, к решению проблемы эмоций, осуществляется, в основном, привлечением вновь открывшихся технических возможностей исследования. Полученные результаты скромно обобщаются простой гипотезой, логика которой, как правило, самоочевидна. “Фундамент” здания науки об эмоциях, заложенный не лучшим образом, невольно заставляет тех, кто изучает эту проблему, ходить общими коридорами, созерцать один и тот же пейзаж из окна и делать выводы, мало отличающиеся друг от друга.
Попробуем рассмотреть появление структур, связанных с эмоциональной сферой, в контексте эволюции нервной системы. В мозге присутствует поле сознания, открытая динамическая система, расположенная между внешней средой и нервной тканью. Вследствие постоянных изменений внешней среды, в эту систему поступает волны энергии, которые изменяют структуру нервной системы и находят выход в активности мышц. Таким образом, энергия внешнего воздействия превращается в механическую энергию, в результате чего достигается равновесие между организмом и внешней средой.
В наиболее общем виде, в нервной системе можно выделить два отдела. Сенсорный мозг, отдел нервной системы, куда поступают потоки импульсов от сенсорных анализаторов (проприоцептивного, зрительного, слухового, вкусового). Моторный мозг, откуда волны избытка энергии переходят в гормональную и мышечную ткани, формируя гомеостаз и программы внешнего поведения организма . Обе части мозга имеют совместные зоны, в которых происходит объединение обоих, динамически отличных систем мозга.
Активность моторной части мозга автоматизирована. Однако организм не может быть полным автоматом в условиях постоянно изменяющейся обстановки. Следовательно, поток импульсов, поступающий через сенсорные анализаторы в мозг, создает в нервной системе избыток энергии, которая не может найти выход в существующих программах поведения. На этот феномен обратил внимание Л.В. Крушинский, в гипотезе об уровне возбудимости мозга, как модификаторе поведения животных (67). Куда деваться энергии? Основная ее масса находит выход в мышечной активности. Биологам хорошо известно, что любой живой организм, в необычных условиях, начинает совершать хаотические, бессмысленные действия. Таким образом:
Избыток энергии формирует целый класс бессмысленных действий, единственной задачей которых является его выход из нервной системы
По законам ассоциирования, если хаотические действия совпадают с определенными образами, то они запоминаются совместно с ними. Следовательно, кратные хаотические движения, ассоциируясь с образами сознания, превращаются в знаки. Теперь, хаотическое движение, знак, воспроизводит в сознании образ, с которым он сопряжен. У животных поворот головы, движения хвоста, поза – выполняют те же функции, что слово у человека. Хаотические движения, принявшие форму знаков, не только выводят избыток энергии из организма, но и оказывают влияние на динамику образов сознания, вырабатывая новые программы поведения, методом проб и ошибок (процесс мышления).

Сенсорные участки моторной сети А и участки сенсорного мозга ВС, образуют совместные зоны АВС, которые управляют теми или иными программами поведения. На определенном этапе развития, гиппокамп и несколько позднее появившаяся лимбическая кора, становиться зоной, где собирается сенсорная информация от всей нейронной сети. С появлением таламуса и новой коры, таких зон становиться две. Гиппокамп, будучи представлен в поле сознания лимбической корой, порождает феномен эмоций.
Представив нервную систему, как динамическую модель, состоящую из двух отделов, попробуем, на примере сенсорного мозга, обратить внимание на некоторые особенности ее эволюции. Нервная система гидры представлена рассеянными элементами, образующими однообразную, сенсорно-двигательную сеть (однослойная нервная система). У червей, нервная система ставиться сложнее. В ней появляются нервные узлы, в которых содержится более совершенная “копия мира”. Основную массу сенсорного мозга гидры и червей, составляют аппараты внешней, контактной рецепции.
Более сложно устроена нервная система насекомых. На базе контактной рецепции в ней формируются рецепторные аппараты, способные воспринимать зрение и слух, новые сенсорные модальности. В нервной системе насекомого, четко просматривается многослойное строение. Возникает феномен цефализации, когда вновь образованный слой нейронной сети, не связан непосредственно с внешней средой и контактируют с ней, через посредников, ранее образовавшейся структуры. Например, мозг перепончатокрылого состоит из трёх отделов: первичный мозг, связанный со зрительными долями, вторичный мозг, воспринимающий рецепцию от органов чувств в антеннах (усики, щупальца) и третичный мозг, который координирует их деятельность (140). Как видим, масса сенсорного мозга прирастает за счет развития его внутренних структур, не имеющих прямого контакта с внешней средой.
Таким образом, по мере эволюции, нервная система превратилось в многослойное образование, где непосредственно контактирует с внешней средой только первый ее слой. Неизбежным следствием такого развития событий, согласно логике противодействия, будет образование “пирамиды”, где элементы нижнего слоя объединяются в верхних слоях, в единое целое.

Сенсорные анализаторы, в процессе формирования многослойной нейронной сети, образуют друг с другом совместные участки. В конечном счете, формируется супермодальность, область, где объединяются все сенсорные модальности.
На схеме расположенной ниже, показан принцип организации сенсорного мозга. Отдельные сенсорные анализаторы образуют совместные участки, которые, в свою очередь, объединяются между собой. (Совместные участки второго слоя, это своеобразные “глаза” внутри нервной системы, но “видят” они лишь то, что происходит в предыдущем слое). Как результат, в нервной системе появляется участок, который является общим для всех видов модальностей. Назовем его “ супермодальность”. По нашим представлениям, областью, соответствующей супермодальности, является гиппокамп и структуры лимбического комплекса, круг Пейпеца (30, 106).

Миндалина (древний моторный мозг), получая информацию от внутренней и внешней сред, окружающих ткани организма, адаптирует его к внешней среде. Гиппокамп и энториальная кора образуют древний сенсорный мозг, который запоминает фон, на котором происходит развертывание той или иной программы поведения, что значительно расширяет адаптационные возможности миндалины. Развитие таламуса и нового сенсорного мозга, еще в большей степени расширяет эти возможности.
На схеме, мозг человека имеет две зоны супермодальности. Первая, более древняя, это гиппокамп, в котором объединяется информация от наружной и внутренней сред, окружающих ткани организма, в единое целое. С появлением второй супермодальности, таламуса и новой коры, гиппокамп получает в ней проекцию (цингулярная и энториальная кора). Именно эти участки коры, участвуя в формировании поля сознания, порождают в нашем сознании феномен эмоций. Вход в новую кору, участок таламуса, связанный с лимбической системой, можно сравнить с “внутренним глазом”, который воспринимает древнюю сенсорную модальность, “запах гиппокампа”. Аналогия с запахом не случайна, так как по своим свойствам, данная сенсорная модальность, более всего напоминает супермодальность (в обонятельной картине мира нет отдельных элементов, предметности, а есть “запах”, суммарная оценка всей внешней среды). По этой причине, потоки импульсов, поступающие в обонятельный мозг, легко соединяются с потоками импульсов в лимбической системе (круг Пейпеца).
Сенсорная информация поступает в мозг через органы чувств. По мере усложнения нервной системы, ее новые слои, не связанные с внешней средой и контактирующие с “центральными концами” периферических анализаторов, образуют совместные участки. В конце концов, появляется зона, где собраны окончания всех анализаторов (гиппокамп). Эволюционный смысл этого явления прост – происходит интеграция всей сенсорной информации в нечто целое, обобщенный образ действительности, содержащий большее количество оттенков, чем любой из первичных сенсорных анализаторов.
На определённом этапе эволюции нервной системы, гиппокамп и окружающие его структуры, выполняли функцию коры мозга. Вся сенсорная информация объединялась тут в нечто целое. Гиппокамп запоминал различные варианты функциональных состояний нижних этажей нервной системы и воспроизводил их в соответствующей обстановке, модулируя активность двигательной части мозга, что проявлялось тем или иным вариантом поведения организма. В случае отсутствия программы поведения, происходил “застой” энергии и искажение существующих программ, т.е. формирование новых программ поведения (процесс мышления).
Вся разнообразная информация от сенсорного мозга, может получить выход, только в одной программе поведения (иными словами, сенсорная информация должна использоваться, как нечто целое). Например, организм, выполняя ту или иную программу поведения, может двигаться только вперед или назад, но не в двух направлениях одновременно. Таким образом, поведение должно подчинятся не одному сенсорному анализатору, а их сумме. “Спор” о приоритете между анализаторами, решается величиной избытка энергии, поступающей в ту или иную сенсорную систему. Уменьшение избытка энергии, говорит об эффективности программы поведения, увеличение, заставляет изменять рисунок поведения, вырабатывать новые программы.
Таким образом, гиппокамп является областью, запоминающей сенсорную информацию, ассоциирующуюся с эффективными программами поведения. Проекция этой области в кору мозга создает ощущение эмоционального фона, который является фактором автоматического извлечения и оценки программ, в соответствии с прошлым опытом, в том числе и приобретенных при участии нового сознания. (Сочетание эмоционального фона со зрительными и слуховыми образами нового сознания, объединяет их с прошлым опытом, делает пригодными для управления поведением человека). Включение содержимого старого сознания в состав нового, в соподчиненном виде, является жизненной необходимостью. Если этого не произойдет, мы потеряем преемственность в развитии отдельных участков нервной системы и возникнет абсурдная ситуация, когда нашим поведением управляют “два водителя”, старая и новая сенсорная модальность. В конечном счете, получаем следующее определение эмоций:
Эмоции, это воспроизведение в сознании всех сенсорных модальностей, как единого целого, наряду с присутствием в нем, первичных сенсорных модальностей в раздельном виде
12. Теоретические вопросы эмоциональной сферы
Самым большим заблуждением, в современной теории эмоций, является представление о том, что они могут быть отрицательными и положительными. Эмоции являются суммой сенсорной информации, сопровождающей конкретные образы сознания. Как любая информация, они получают оценку лишь в контексте практического применения, то есть поведения. Образ сознания всегда имеет своеобразный маркер, определенный эмоциональный фон, который может восприниматься как положительным, так и отрицательным. Так, демонстрация стрелкового оружия вызывает положительные эмоции у большинства мужчин и отрицательные, у большинства женщин. Как видим, определение “положительный” или “отрицательный” не есть качественная характеристика образа сознания и соответствующей эмоции. Эмоциональная оценка одного и того же явления, может диаметрально изменяться, в зависимости от контекста ситуации. Например, в гостях у товарища мы ощущаем “атмосферу его дома” - цельный эмоциональный образ, имеющий положительный оттенок. Предположим, что товарищ совершил, по отношению к нам, неблаговидный поступок, который нас поссорил. Теперь эмоциональный образ “атмосферы дома” приобретает, в нашем восприятии, отрицательный оттенок. Или другой пример. Приступая к решению сложной задачи, мы ощущаем эмоциональный подъем. Однако если задачу решить не удается, эмоциональный фон, в нашем сознании, приобретает неприятный оттенок.
Таким образом, эмоциональный фон образа не может быть изначально положительным или отрицательным. Феномен знака эмоции надо объяснять иначе. Образы нашего сознания, это следствие поступления энергии внешней среды в нервную систему. Если энергия без проблем преобразуется в процесс поведения, то образы в сознании не появляются, а мы говорим об автоматизированных действиях. В ситуации, когда автоматические навыки недостаточны, для уравновешивания изменений окружающей обстановки, в нервной системе происходит застой энергии, активация сознания, процесса мышления и изменение программ поведения. Длительное существование в нервной системе неэффективной программы поведения, сопровождается застоем энергии и отрицательным эмоциональным фоном. Найденное решение задачи придает тому же фону положительный знак.
Следовательно, положительная или отрицательная оценка образа зависит не от качества сопровождающей его эмоции, а от того, как долго он присутствует в нашем сознании избыток энергии, обусловивший появление эмоционального фона. Например, если мы приехали на отдых, сменив место жительства, то новая обстановка будет восприниматься нами положительно лишь определенное время. Со временем, мы станем к ней равнодушны, перестанем замечать, а возможно, даже, станем испытывать к ней отрицательные чувства. Оценка “плюс” или “минус”, это не оценка качества сенсорного образа, а ощущение того, что можно делать с этим образом (то есть характеристика программы поведения, возможности вывода энергии из нервной системы и достижения равновесия с окружающим миром). Быстрое исчезновение сенсорных образов из сознания, при наличии соответствующей программы поведения, даёт нам, представление о чем-то положительном. Длительное присутствие образа в сознании придает, тому же эмоциональному фону, отрицательный оттенок.

На графике показано, что величина избытка энергии и длительность его существования, основные параметры, определяющие качество эмоций.
Эмоции, в своей сущности, это избыток энергии в нервной системе. Отсюда следует, что они отрицательны, по своей природе. У ребенка, любое изменение внешней среды вызывает дискомфорт, отрицательные эмоции и требование “вернуть все как было”. Удовлетворение потребностей (прием пищи) переводит отрицательный эмоциональный фон, в эмоциональный фон положительный (104). Таким образом, положительный знак эмоции, возникает в случае эффективности поведения, как феномен снижения избытка энергии в нервной системе. Знак “плюс” и “минус” свидетельствует об эффективности программы действий, а не о каком-то особом содержании сенсорной информации (это автоматическая оценка сознанием своих возможностей). Следовательно:
Эмоциональное ощущение, это признак появления в сознании избытка энергии, что в свою очередь определяется эффективностью программы поведения
Отрицательный знак эмоции определяется нарастанием избытка энергии в нервной системе, а положительный, его убыванием
Попробуем применить вышеизложенное представление об эмоциях, к современным оценкам этого явления. А.С. Батуев, сравнивая эмоции и мотивации, пишет (18):
“До сих пор нет четкого мнения о том, различаются эти два субъективных состояния организма или отражают просто разные оттенки одного и того же процесса”
Точки зрения, что эмоции это проявление мотивации, придерживались Ю.М. Конорски им П.К. Анохин. К.В. Судаков, предложил следующее определение:
“ Мотивация представляет собой возникающее под влиянием первичных изменений во внутренней среде эмоционально окрашенные состояния организма, характеризующиеся избирательными активирующими влияниями специальных подкорковых аппаратов на кору головного мозга и другие его отделы и направляющие поведение животного на удовлетворение исходной потребности”
Как видим, нет понимания, что эмоция – это сумма сенсорной информации. Если происходят изменения внутренней среды организма, мы, к примеру, имеем чувство голода, которое активирует пищевое поведение. Если изменения касаются внешней среды, например появление чего-то необычного, возникает чувство любопытства и исследовательское поведение. Эмоции – это смесь всех сенсорных ощущений, а мотивация, та их часть, появление которой определяется изменениями внутренней среды организма.
Биологическая концепция эмоций, предложенная Ч. Дарвином, является не их объяснением, а остроумной классификацией, в основу которой положены особенности мимики животных. Мимика – это язык древнего сознания. До того как появились “речевые движения”, отражающие состояния нового сознания, существовали движения мимические. Набор этих “двигательных штаммов” отражает настроение, те или иные разновидности основные разновидности состояний сенсорной сферы.
П.К. Анохин характеризует эмоции верно, но неясно. По его мнению, эмоции, это субъективные ощущения, позволяющие быстро оценить внутренние потребности, действие внешних факторов и результаты поведения. То есть, эмоции – это сумма сенсорной информации, автоматически придающая сознанию определенный вектор движения, с возможным внешним проявлением в виде реализации потенциальных программ действий. Наша гипотеза предполагает, что избыток энергии, следствие нарушения равновесия между организмом и внешней средой, активирует сознание новое и древнее (эмоции). Эмоции – это сигнал о появлении избытка энергии в нервной системе, который, приводя в движение содержание сознания, активирует или создает нейтрализующие этот избыток программы поведения. По мнению П.К. Анохина, любая потребность сопровождается эмоциональным переживанием отрицательного характера, а ее удовлетворение сопровождается положительным эмоциональным ощущением. Как видим, знак эмоции зависит от эффективности программы поведения (рост избытка энергии в нервной системе придает эмоции отрицательный оттенок, а его снижение, меняет отрицательный знак эмоции на знак положительный).
Таламическая теория эмоций В. Кеннона и В. Барда, связывает проявление эмоций с активацией таламичсеких структур. По нашему мнению, данная теория путает причину и следствие. Эмоции возникают не в результате снижения тормозящего влияния коры на таламус, а в результате нарушения равновесия в нервной системе и появления в ней избытка энергии.
Периферическая теория эмоций Джеймса-Ланге, согласно которой эмоции отражают изменения в тканях организма, есть правильное, но одностороннее понимание проблемы (113). Тоже можно сказать о лимбической теории эмоций, которая связывает их появление с глубинными отделами мозга, выполнявшими, на определенной ступени эволюции, функцию сознания (в появлении эмоций участвует вся сенсорная система мозга, сенсорной информации без эмоций не существует).
Представительство лимбической системы в коре головного мозга человека (поясная извилина, грушевидная извилина), находиться в соподчиненном статусе по отношению к сознанию. Удаление ключевых образований лимбической системы создает дефект в эмоциональном фоне сознания, нарушая программы поведения. Лимбическая система не только “висцеральный мозг”, как ее характеризует П. Мак-Лин, а полноценный мозг, конвергирующий входы от всех сенсорный модальностей, на уровне ствола мозга. В тоже время, П. Мак-Лин остроумно замечает, что различие между новой корой и лимбической системой такое же, как между, “мы чувствуем” и “мы знаем”, с чем невозможно не согласиться.
В рамках излагаемого материала, очевидно, что возбуждение ретикулярной формации сопровождается ростом избытка энергии в мозге, с последующим изменением эмоционального фона (активирующая теория эмоций Д.Б. Линдслея). Однако надо понимать, речь может идти только о модуляции уже существующего эмоционального ощущения.
Вопрос о том, где локализован участок мозга, отвечающий за эмоциональное ощущение, не совсем корректен. Эмоции, по определению, не могут быть локализованы, так как в их образовании, равно как и в образовании сознания, участвуют все афферентные потоки мозга. Правильней поднимать вопрос: “Как повреждение той или иной области мозга оказывает влияние на качество данной, конкретной эмоции?” Наибольший дефект эмоциональной сфере наносит повреждение образований лимбического комплекса, где высока концентрация сенсорной информации. В отношении центров “удовольствия” и “неудовольствия” необходимо заметить: раздражение участков нервной системы, которые способны вывести поступающую в них энергию в силу своего анатомического положения или функционального состояния, сопровождается ощущением положительного эмоционального фона, поэтому данный участок будет отнесены экспериментатором к центру удовольствия. Если выход энергии из раздражаемого участка затруднен, то эмоциональный фон будет отрицательным и заключение экспериментатора будет соответствующим.
Интересна потребностно-информационная теория эмоций В.П. Симонова, которая рассматривает проблему эмоций в контексте поведения. Автор предлагает формулу эмоций (114):
Э = П (ИН – ИС)
Э – эмоция, ее сила, качество, знак. П – сила и качество актуальной потребности. ИН – информационные средства необходимые для ее удовлетворения. ИС – средства имеющиеся в наличии. Как видим, ИН и ИС, это программы поведения и если они недостаточны, эмоция отрицательна.
Наше понимание сущности эмоций позволяет несколько иначе представить данную формулу:
Э = ИЭ (ПП – ПН)
Э – эмоция. ИЭ – избыток энергии в нервной системе. ПП – имеющиеся в наличии программы поведения (потенциальные). ПН – программы поведения, необходимые для вывода избытка энергии из нервной системы.
Как видим, если программ избыточное количество, достаточное для быстрой выработки новых программ поведения, избыток энергии без задержек выводиться из нервной системы, поэтому знак эмоции положительный. Если программное обеспечение недостаточное, знак эмоции становиться отрицательным. Наконец, если программа поведения совпадает с внешним воздействием, то действия контролируются на бессознательном уровне и эмоция равна нулю (избыток энергии до сознания не доходит).
Направленность процессов в нервной системе определяется, с одной стороны, изменениями внешней среды, а с другой, противодействием живой системы этим воздействиям (то есть, вектор поведения организма определяется не только потребностями, но и изменениями внешней среды). Поэтому правильнее говорить, что эмоциональные процессы выражают не только потребности, но и воздействия окружающей среды, а степень участия в их формировании этих процессов не есть величина постоянная.
Предлагаемая гипотеза эмоций объясняет изменения сознания, наступающие при двустороннем удалении гиппокампа (такие больные не могут усваивать текущую информацию, при полной сохранности прошлого опыта и интеллекта). Как следует из логики излагаемого материала, при удалении гиппокампа теряется механизм интеграции афферентных потоков, поступающих через отдельные сенсорные анализаторы, поэтому информация, поступившая через один из них, становится доминантной, заполняет все сознание и определяет текущее поведение (для больных с данным дефектом характерно, что их поведение управляется последней, поступившей в сознание информацией). Кроме того, теряется механизм запоминания текущей информации, так как образ сознания не получает своей “тени”, эмоционального фона и не может включиться в прошлый опыт (воспринимается как нечто совсем новое, не имеющее аналогов в архивах памяти).
До сих пор мы рассматривали процесс формирования эмоционального ощущения при активном участии мира вокруг нас. Однако его появление в сознании может быть следствием внутренней активности нашего Я, без участия внешней среды. Например, эмоциональные опущения во время сна или при творческой работе, в процессе мышления, когда движение образов сознания происходит при минимальном участии внешней среды. Эмоциональные ощущения, возникающие в подобной ситуации, можно выделить в отдельный класс, который будет соответствовать понятию “эстетические ощущения”. Таким образом:
Чувства, эстетические ощущения – это эмоциональный фон, возникающий в результате процесса мышления.
С данных позиций не так уж трудно понять, что представляет собой интуиция. Эмоциональный фон, смесь сенсорных модальностей, учитывает все доступные восприятию особенности внешней обстановки. Его формирование происходит мгновенно, как процесс восстановления равновесия между процессами, протекающими в мозге и изменившейся внешней обстановкой. Одновременно из памяти извлекаются программы поведения, соответствующие прошлому опыту. Решение теоретической задачи сопровождается эмоциональным фоном, который автоматически включает программы внутреннего поведения (мышления), близкие к поставленной задаче (соответствующие программы внутреннего поведения потенциально находятся в нашей памяти). В тех случаях, когда задача не имеет готового решения, происходит многократное извлечение из памяти неэффективных алгоритмов, что, в конечном счете, приводит к искажению самих алгоритмов и последовательности их воспроизведения. Когда процесс искажения, в силу случайных сочетаний, происходит быстро, мы говорим об интуиции, не замечая за ней длительный процесс накопления программ внутреннего поведения, бывших до этой поры неэффективным.
В заключение попробуем разобраться, каким образом музыка воздействует на эмоциональную сферу. Лимбическая система – это “склад эмоций”. Рядом с ней, в височной доле, расположена первичная и вторичная слуховая кора. Слуховые образы являются чем-то средним между двумя крайностями, образами обонятельными и зрительными (в том плане, что сумма слуховых образов воспринимается как нечто целое и это целое практически не имеет топологии в пространстве). В процессе эволюции, даже на уровне новой коры, совместные участки, через которые возможен обмен энергии между слуховой и лимбической корой, хорошо развиты.
При исполнении музыкального произведения, структура музыкальной фразы начинается с небольшого фонического разнообразия. Далее, силовая и структурная сторона звучания нарастает, пока, в конечном счете, не наступает уменьшение параметров звучания. Что же происходит, в этот момент, в нервной системе? По мере усиления звукового воздействия в ней растет количество энергии, выход которой определяется логикой динамики музыкальной фразы (резкий спад уровня энергии в мозге, по окончании музыкальной фразы, способствует появлению положительного эмоционального фона). Таким образом, при прослушивании музыки, поток слуховых образов, сопровождаемый значительным эмоциональным фоном, становится основным фактором, модулирующим эмоциональную сферу. Если принять во внимание удивительное многообразие музыкальных фраз, которые волнами следуют одна за другой, порождая необычные эмоциональные ощущения высокой амплитуды, становится понятно, откуда берется сила музыкального воздействия на слушателя.
13. Функциональная ассиметрия мозга
По мнению Дейвидсон и Геллер, знак эмоций зависит от соотношения активности левой и правой фронтальной коры (113). Этому и ряду других фактов, касающихся функциональной ассиметрии мозга, можно найти несколько другое объяснение.
Как обсуждалось ранее, наряду с циклической, автоматической, формой движения, в нервной системе существует линейная, хаотическая форма движения, представленная ориентировочным рефлексом. По мере эволюции, эффективные ориентировочные действия запоминаются, что приводит к их усложнению и упорядочиванию. Упорядоченный и усложненный ориентировочный рефлекс проявляется способностью сознания, при смене своих состояний, действовать по определенным правилам, которые все больше начинает влиять на динамику процессов в нервной системе и, в конечном счете, начинает ею управлять (формируется логика мыслительных процессов).
Движение внутри динамической системы подразумевает определенную последовательность событий, без которой отдельные ее части вступят в противоречие друг с другом. Ориентировочный рефлекс, как фактор управления поведением, не может формироваться в двух полушариях мозга одновременно, поэтому, закрепившись по ходу эволюции в одном из полушарий, он вынужден в нем оставаться и в дальнейшем. Управление поведением не может осуществляться одновременно двумя участками мозга, как не имеет смысла управление самолетом двумя пилотами, которые не считаются друг с другом. Таким образом, развитие доминантной группы мышц и связанного с ней правого или левого полушария, определяется случайными факторами внешней обстановки и закрепляется по ходу развития нервной системы.
Если в левом полушарии формируются структуры, накапливающие опыт ориентировочной деятельности, то в правом полушарии запоминается фон, на котором он происходит. Фактически, кора правого полушария, становится последней инстанцией, верхним слоем нейронной сети, по отношению к левому полушарию. Благодаря участию симметричных участков коры правого полушария, программы ориентировочной деятельности, записанные в левом полушарии, учитывает большее количество оттенков внешней среды и соответственно их уравновешивает.
На схеме видно, пусковым механизмом появления функциональной ассиметрии правого и левого полушарий является первичное поступление энергии в левое полушарие, сложившиеся в процессе эволюции, с последующим формированием в нем структуры ориентировочного рефлекса (при этом доминантность усиливается по мере развития фронтальной коры).

Энергия внешней среды поступает в доминантное полушарие, где происходит, в процессе мышления, формирование программ поведения. Поступление энергии в правое полушарие, формирует дополнительный блок памяти, благодаря которому расширяются возможности энграммы.
В 1970 году, Sperry, изучая мозг с перерезанным мозолистым телом, обнаружил характерный когнитивный дефект. В проведенных им экспериментах больные получали следующее задание: в соответствии с предъявленным рисунком предмета или его словесным обозначением найти на ощупь данный предмет за перегородкой из числа нескольких других. Проверялось выполнение данного задания при изолированном предъявлении изображения только в правое или левое полушарие. Оказалось, что в обоих случаях больной успешно выполнял задание. Но если изображение поступало в левое полушарие, то больной мог дать словесный отчет о своих действиях. В тоже время при предъявлении изображения в правое полушарие больной не мог сказать, что и почему он сделал. Таким образом, он в этом случае как бы не сознавал своих действий.
Вышеизложенная точка зрения помогает объяснить этот феномен. В левом полушарии, путем проб и ошибок, формируется новая программа поведения. Правое полушарие пассивно запоминает уже готовую программу и фон, на котором она происходит. Таким образом, в правом полушарии существуют уже готовые программы поведения, которые активируются на предъявление стимула (когда память сформирована, она автоматически воспроизводит консолидированную в ней информацию, в соответствии с текущей обстановкой). Выполнение задания, при поступлении информации исключительно в правое полушарие, активизирует готовые программы поведения, а последовательность выполняемых действий не фиксируется сознанием. Подобная задача в левом полушарии активизирует ориентировочный рефлекс и, по сути дела, решается заново.
Экспериментально установленный факт эмоциональной ассиметрии правого и левого полушарий, закладывается при развитии их доминантности. Задача, стоящая перед сознанием, активно решается структурами левого полушария, что приводит к уменьшению избытка энергии в этой части мозга. Если задачу решить не удается, застой энергии больше заметен в правом полушарии, так как оно пассивно наблюдает за действиями левого полушария и не использует поступающий в него избыток энергии.
Асимметричность поведения, древнейшее свойство любой активной биологической системы. В тоже время, доминантность одного из полушарий, следствие ассиметричной активности организма во внешней среде, закрепляется филогенетически только при определенном уровне развития нервной системы.
14. Интеграционные процессы мозга
Многие явления внутри нервной системы остаются непонятными по той причине, что мы не можем осознать принципы организации процессов, обеспечивающих сознание человека. Попробуем исправить положение. Будем исходить из того, что мир вокруг нас является материей, которую, в виду особенностей ее восприятия, мы предлагаем рассматривать, как линейную и циклическую формы движения. Представление о сознании человека, как особой форме материи, образуемой циклическими потоками импульсов в нервной системе, помогает иначе расценивать происходящие в мозге явления.
С этой точки зрения, динамическая система сознания, равно как и любая другая динамическая система, будет стремиться к равновесию с внешней средой. Потоки импульсов на поверхности нейронной сети образуют своеобразную оболочку, которая является посредником между окружающей средой и тканями организма. В виду изменчивости внешней среды, равновесие с ней постоянно нарушается и энергия, через многочисленные рецепторные аппараты, поступает в поле сознания. Избыток энергии уравновешивается перестройкой всей системы кольцевых потоков импульсов и суммарной мембраны нейронов, на которую эти потоки “опираются” (мембрана обладает определенной пластичностью и, изменяя свою структуру, способна уравновешивать множество состояний внешней среды, что составляет суть феномена памяти в нервной системе).

Поток импульсов на поверхности нейронной сети, является буфером между суммарной мембраной нейронов и внешней средой. Оболочка из потоков импульсов имеет контакт с внешней средой только в области рецепторных аппаратов, а в остальной части она соприкасается с мембраной нейронной сети, которая и заключает в себе прошлый опыт существования организма.
Многие исследователи догадывались, что мозг следует рассматривать, как самоорганизующуюся систему, однако отсутствие философской концепции не позволяло развивать возникшие предположения. Еще Р.Декарт остроумно проводил аналогию между происходящими в нервной системе процессами и жидкостью, заключенной в замкнутом пространстве. С этих позиций оценивал нервную систему и А.А. Ухтомский. Развивая представления о доминанте и условном рефлексе, он писал (18):
“Связь исполнительного органа с новыми рецептивными поводами может закладываться, так сказать, случайно, вследствие того, что работа данного исполнительного органа имела случаи – более или менее совпадать во времени с данной дальней рецепцией. Кора бдительно примечает такие совпадения, закладывает по их поводу новые связи, сначала временные, а затем могущие закрепиться в качестве опыта и фонда, который будет использован животными для дальнейшей жизни… перед нами новая рефлекторная дуга в процессе своего закладывания и дальнейшего закрепления”
А.А. Ухтомский выделяет три стадии доминанты: безразличное привлечение к себе любой рецепции, вылавливание из нее биологически значимой и закрепление доминанты за ключевыми внешними воздействиями. Еще ближе к пониманию сущности доминанты подходит А.С. Батуев (в своих комментариях к работам А.А. Ухтомского):
“… за каждой естественной доминантой кроется возбуждение целого созвездия (констелляции) центров, своеобразная циклическая машина, функциональный орган, образующийся по ходу самой реакции”
Как видим, явно просматривается тенденция представить деятельность мозга, как системы, которая стремиться к равновесию с окружающей ее средой. В тоже время, проблема уходит из компетенции нейрофизиологии в сферу интересов философии, что создает определенные трудности для ее решения. Доминанта, это участие определенных участков нервной системы в уравновешивании изменившихся условий внешней среды. Однако, фиксируя очаги возбуждения, мы забывает о том, что участки торможения не менее важны для формирования равновесия между полем сознания и окружающей средой. Оболочка из циклических потоков импульсов реагирует на изменения среды не по частям, а как единое целое. Таким образом, доминанта А.А. Ухтомского, это реакция поля сознания на изменения внешней среды, с точки зрения возбуждения. Система импульсов сознания, воспринимая избыток энергии, входит в новые соотношения с мембраной нейронов, вызывая ее функциональную и органическую перестройку, что позволяет говорить о стадиях доминанты. Стойкие изменения мембраны нейронов совпадают с представлением о закреплении доминанты (третья стадия доминанты, это новый уровень равновесия с внешней средой). Изложение материала в контексте циклической формы движения устраняет некоторые иллюзии, которые тормозят понимание проблемы.
Рефлекс не есть изолированная реакция организма на определенные факторы окружающей среды, это субъективное восприятия общего вектора движения, возникающего в нервной системе в ответ на ее изменения (вероятно, рефлекторную дугу можно проследить не только с точки зрения возбуждения, но и с точки зрения торможения). Искусство экспериментатора направленно, помимо его воли, не столько на объяснение сути рефлекса, сколько на способы получить возможность им управлять (именно это является критерием оценки результата исследования).
Ассоциирование, совместное закрепление ряда факторов внешнего воздействия между собой в памяти субъекта, не связано с появлением “мифических связей” между определенными зонами мозга. Оболочка из потоков импульсов, покрывающая нейроны рефлекторной сети, заключена в “жесткие рамки” представленные с одной стороны свойствами самой мембраны нейронов, а с другой, определенными параметрами внешней среды. Следовательно, сложившееся равновесие с внешней средой задает определенные параметры, как оболочке из потоков импульсов, так и мембране нейронов. Как обсуждалось выше, в равновесных системах можно говорить о феномене заполнения, который гораздо глубже объясняет процессы ассоциирования в нервной системе.

На схеме, группа из девяти нервных узлов, символизирующих нервную систему, заключена в определенные параметры внешней среды. В каждом узле сохраняется определенное сочетание активности составляющих его нейронов, обозначенных как узор информации. Нервный узел 5 поврежден, поэтому в нем отсутствует узор информации.
На схеме представлена модель нервной системы, где существует определенное сочетание активности нейронов, поддерживаемое кольцевыми потоками импульсов. Предположим нервный узел “5” будет поврежден. Учитывая, что он заключен в жесткую оправу динамических процессов, в процессе восстановления его структуры в нем может появиться только узор информации похожий на “УИ5”. Феномен заполнения объясняет “феномен ампутированных”, ложные воспоминания, реализацию двигательных программ разными средствами без предварительного обучения и так далее. Ассоциирование обеспечивается функциональной перестройкой всего мозга, при адаптации к изменившимся условиям, где ассоциируемые факторы включаются в общую динамическую систему и в дальнейшем воспроизводятся, как элементы этой системы, для повторного уравновешивания параметров внешней среды. Именно таким образом можно объяснить парадоксальное высказывание К.С. Лешли о том, что памяти нигде нет, но в то же время, она всюду (106). Следовательно:
Система кольцевых потоков импульсов в нейронной сети отражает все параметры внешней среды, существующие в данный момент времени, способствуя из совместному запоминанию и воспроизведению
Еще одним заблуждением, которое постепенно изживается, по мере накопления экспериментального материала, является представление о возбуждении и торможении в нервной системе, как о двух полноценных системах управления. Торможение должно рассматриваться в контексте пластичности нервной системы. Потоки энергии, обеспечивающие возбудительный процесс в нервной системе, образуют циклическую форму движения на мембранах нейронов, изменяя последние и формируя основные проводящие пути, которые, в свою очередь, активируют соответствующие ткани организма и отражают внешнее воздействие. Поскольку пропускная способность нервных путей ограничена, формируются дополнительные барьеры противодействия: усложнение участка мембраны, контактирующего с внешней средой (специализированные рецепторы отфильтровывают часть энергии среды), увеличение массы узла, формирование вставочных нейронов, развитие нейропиля, изменение свойств синаптической мембраны, в плане ее пропускной способности (габитуация или сенситизация) и так далее.
Один из видов пластичности нервной системы, проявляется в изменении свойств медиаторов, которые вырабатывают окончания нейронов. В местах, где возбуждение, как фактор, обеспечивающий выход энергии из нервной системы, не дает эффекта, может оказаться полезным снижение активности каналов проведения импульсов по нейронной сети. Таким образом, в синапсах, которые вырабатывают возбуждающий медиатор, может произойти искажение его образования. Окончания нейронов, усваивая поступившую энергию, выполняют определенную работу, вне зависимости от того, какого качества медиатор ими вырабатывается, поэтому вставочные нейроны, внутри нервной системы, могут вырабатывать самые разнообразные медиаторы, если появление последних способствует более эффективному достижению равновесия между организмом и средой
Поскольку одностороннее развитие возбудительных процессов не может обеспечить равновесие нервной системы с внешней средой, часть существующих в ней потоков импульсов, перестраивается на участие в тормозных реакциях, что обеспечивает более эффективную утилизацию избытка энергии (для стабильного существования системы необходимо не только возбуждение, движение вперед, но и торможение, движение назад). Таким образом, энергия внешнего воздействия способствует формированию в нейронной сети многочисленных барьеров противодействия, в которых участвуют медиаторы самого различного свойства. Важно отметить, что все разнообразие медиаторов в нервной системе объяснимо, с точки зрения ее развития, как самоорганизующейся системы. Таким образом:
Торможение – одна из форм противодействия внешней среде, в основе которой лежит видоизмененное возбуждение
Возбуждение обеспечивает существование кольцевых потоков импульсов в нейронной сети, а торможение снижает энергию самого кольца, адаптируя всю систему импульсов к изменившимся условиям
15. Принципы организации коры мозга
Представление о циклической форме движения заставляет рассматривать процессы, происходящие в нервной системе, как множество кольцевых потоков импульсов, причем все движение, на мембранах нейронов, имеет иерархическую организацию. Над одной системой колец, по мере развития рефлекторной сети, надстраивается другая, малые кольца, объединяясь между собой, образуют большие и так далее. Вся движущаяся система принимает энергию от внешней среды и отдает ее обратно, через ткани организма. Образы в нашем сознании не что иное, как круговое движение импульсов на суммарной мембране нейронов. Циклический характер движения в нервной системе порождает представление об обратной связи.

Избыток энергии формирует центральный слой коры нервный узел, “развернутый на плоскости”. Рядом с ним возникает слой обратной связи, который обеспечивает кольцевое движение импульсов с участием нервных узлов ствола. Отводящий слой образует такое же кольцо с соседними участками коры. Количество дополнительных слоев определяется величиной избытка энергии, в данной области мозга и обеспечивает расширение возможностей запоминания объема информации.
Кора мозга – наиболее молодой участок рефлекторной сети. Каждый ее слой представляет собой развернутый на плоскости нервный узел, сохраняющий топографические особенности своего периферического прототипа. По логике вещей, в слоях коры должен быть “узел” принимающий узор информации, “узел” отдающий его обратно и “узел” направляющий его в другие, совместные участки нервной системы. Поэтому, основные принципы организации коры мозга следует схематически представить следующим образом. Нумерация слоев коры от первого до шестого и более, не совсем верна, с точки зрения ее функциональной организации. Надо различать основные слои (которые мы условно обозначим как “центральный”, “обратной связи” и “отводящий”) и вспомогательные, “модулирующие”, количество которых определяется функциональной нагрузкой на данный сенсорный анализатор.

Принципиальная схема движения кольцевых потоков импульсов в коре мозга, где ЦС – центральный слой, СОС – слой обратной связи, ОС – отводящий слой.
Анфилада блоков нервных узлов образует первичную, вторичную и третичную зоны коры. Благодаря движению паттернов импульсов по этим структурам, энергия внешнего воздействия рассеивается, что является своеобразным барьером противодействия внешней среде, со стороны сенсорной части мозга. Нет необходимости искать выход из коры к мифическому адресату, обязанному "принять решение". Энергия внешней среды, проходя через ткани организма, способствует становлению равновесия между организмом и средой. "Раздутая" на блоки сенсорная кора мозга, частично нейтрализует избыток энергии, придавая ему форму образов нашего сознания, а частично отдает его в моторный мозг, перестраивая кольца движения импульсов, изменяющие рисунок мышечной активности и повышая уровень адаптации организма в среде.
Связь между различными участками нервной системы возникает в процессе передвижения избытка энергии от одного кольцевого потока импульсов к другому. Волны движения, проходящие через рецепторное поле, отражаются от участков суммарной мембраны нейронов, взаимодействуют друг с другом (в результате изменяется уровень пульсации многих, часто значительно удаленных друг от друга нейронов) и, приняв соответствующую форму, уравновешивают процессы внешней среды.

На схеме показан третичный узел, где, благодаря общему нейропилю, происходит объединение двух вторичных зон коры мозга, что позволяет их рассматривать, как единое, функциональное образование. В тоже время, подобное объединение позволяет вторичным зонам не смешиваться друг с другом и сохранять определенную самостоятельность.
Третичные зоны коры мозга имеют связь с несколькими, нижерасположенными участками нейронной сети, имеющими самостоятельные сенсорные входы. Таким образом, третичный участок сети является узлом, в котором оканчиваются и сосуществуют несколько кольцевых потоков импульсов (например, от двух сенсорных анализаторов). Третичный узел не есть механическое соединение нескольких нервных узлов. Благодаря нейропилю, общему для всего узла, избыток энергии отражается от суммарной мембраны всех нейронов данного объединения и формируется совместный узор информации нейропиля третичного узла. Теперь узор информации, пришедший в один из узлов, является достаточным фактором для активации общего узора информации, записанного в памяти мембраны данного узла. Данный процесс, известный как ассоциирование, способствует выработке совместных узоров информации, которые модулируют форму избытка энергии, волны которого периодически поступают в нервную систему. Следовательно:
Участки коры, в которых объединяются несколько сенсорных анализаторов, уравновешивают избыток энергии, поступивший по одному из каналов, делая возможным его выход через остальные сенсорные каналы
Как видим, благодаря общему нейропилю и перестройке совместной мембраны нейронов третичного узла, становиться возможным превращение звука в слово (моторной информации в сенсорную информацию и наоборот), а также появлению образов сознания, в которых участвуют несколько сенсорных анализаторов. Происходит значительное расширение возможностей, как процедурной, так и декларативной памяти. Одни и те же программы поведения, используются в сочетании с новыми образами сознания, создаются более благоприятные условия, для выработки новых программ поведения, намного увеличивается количество оттенков образов сознания.

Избыток энергии, поступая в нервную систему, изменяет уровень энергии в кольце движения импульсов, что передается следующим кольцам, порождая волновое движение. Поскольку потоки импульсов ограничены, с одной стороны, мембранами нейронов, а с другой, параметрами внешней среды, параллельно с выходом энергии из нейронной сети, в виде активности тканей организма, видоизменяются и свойства мембраны нейронов и уровень его активности.
16. Зрительный анализатор
Через зрительный анализатор проходит, по некоторым данным, до 90% всей сенсорной информации. Несмотря на то, что он является одной из наиболее изученных областей мозга, до сих пор нет ясного представления о том, как он функционирует. В данной главе мы попробуем исходить из представлений, что зрительный анализатор является самоорганизующейся системой, стремящейся прийти к равновесию с окружающей его средой. В самом начале изучения анатомии глазного яблока, многих исследователей приводило в недоумение совершенство его структуры. Поневоле напрашивалась мысль о существовании творца, который, создавая глазное яблоко, предвидел все особенности его дальнейшей эксплуатации. Все выглядит вполне объяснимо, если рассматривать эволюцию зрительного анализатора в контексте самоорганизующейся системы. Глазное яблоко, есть продукт длительного процесса противодействия рецепторного аппарата внешней среде, а сложность его анатомической структуры находиться в прямой зависимости от количества проходящей через него энергии и уровня равновесия с окружающей средой.

Средний фоторецептор, возбуждаясь сильнее, чем его соседи, тормозит их активность посредством горизонтальной клетки, которая является дополнительным барьером на пути внешнего воздействия. Ту же роль играет аквамариновая клетка, утилизирующая избыток энергии, поступающий по системе обратной связи из верхних слоев зрительного анализатора.
Попробуем рассмотреть структуру зрительного анализатора, как структуры противодействия внешней среде. Фоторецепторы являются первым фильтром на пути светового потока. Процесс противодействия предусматривает уравновешивание определенных факторов внешнего воздействия. Это неизбежно приводит к тому, что в фоторецепторах будет присутствовать несколько упрощенная динамическая копия светового потока и объектов, от которых он отражается. Разнообразие фоторецепторов по свойствам и месту расположения на сетчатке обусловлено, с одной стороны, неравномерностью светового потока, а с другой, возможностью дальнейшего выхода энергии поступившей в нервную систему (понятие биологической значимости несколько вуалирует сущность происходящих процессов).
Структура сетчатки имеет стандартные механизмы противодействия внешней среде. Как показано на схеме, сетчатка является своеобразным фильтром, отсекающим часть внешних воздействий, но сохраняющим логику их динамики, в проходящем потоке импульсов. Сетчатка нейтрализует фоновую освещенность, пропуская только локальные колебания светового потока, выделяющиеся на общем фоне (через нее “проскакивают” точечные воздействия, которым противостоят единичные рецепторы). Энергия, воспринятая одним из группы фоторецепторов, на какое-то время становиться выше уровня локального, тормозного противодействия горизонтальной клетки (активация всех фоторецепторов, связанных с данной горизонтальной клеткой, повышает уровень ее противодействия, поэтому импульсы не поступают в сенсорный анализатор). Можно сказать, несколько упростив ситуацию, что неподвижный глаз, в момент локального изменения освещенности, пропускает "точки" и "группы точек" (линии), которые внутри зрительного анализатора создают узор информации, уравновешивающий определенные параметры изменившейся освещенности среды.
После того, как первые слои сетчатки уравновешивают световой поток, в нейронной сети происходит перестройка кольцевых потоков импульсов, что соответствует формированию в них образа внешнего мира в зрительном анализаторе. Необходимо подчеркнуть, что фоторецепторы реагируют не на качество объектов внешнего мира, а восстанавливают равновесие со световым потоком, поэтому им не важно, происходит усиление или снижение светового воздействия (on-off-реакция).
Избыток энергии поступает в сетчатку не только со стороны окружающей среды, но и со стороны нервной системы. Локальное изменение узора информации (паттерна импульсов) в коре, меняет свойства кольцевого потока. Волна изменений распространяется, по структурам зрительного анализатора, во всех направлениях, в том числе от центра на периферию. Достигнув ганглиозных клеток, обратная волна энергии перестраивает их активность (формирует новый узор информации). Таким образом, избыток энергии, приносимый возвратной волной энергии, порождаемой перестройкой активности нейронов зрительной коры, частично изменяет активность ганглиозных клеток, а частично нейтрализуется аквамариновыми клетками, которые являются функциональным аналогом горизонтальных клеток, но ориентированным не на противодействие внешней энергии, а на противодействие энергии поступающей по обратной связи.
Из сетчатки волна избытка энергии и соответствующий ей узор информации переходит в коленчатое тело, которое, на определенном этапе эволюции, образовало связь с моторной частью нейронной сети, обеспечившей вывод избытка энергии.

Энергия внешнего воздействия, поступившая из сетчатки в коленчатое тело, находит выход через верхние бугорки четверохолмия, активируя движение глазодвигательных мышц.
В работе, посвященной проблемам сна, Н. Клейтман писал (139):
“Вполне логично предположить, что деятельность глазодвигательных мышц, а так же мышц век, может служить весьма чувствительным индикатором изменений, происходящих в нервной системе во время цикла сна и бодрствования. Обширность областей коры, влияющих на движение глаз, наличие путей от вестибулярного аппарата к глазодвигательным ядрам, с одной стороны, и относительно малое число волокон, иннервирующих мышцы глаза – с другой, составляют, по меньшей мере, количественную основу возможной связи движения глаз с общей активностью центральной нервной системы. А анатомическая близость ядер глазодвигательных нервов к путям, участвующим в поддержании бодрствования указывает на возможность более специфических взаимоотношений”
Эволюция глазодвигательной системы , является самоорганизующимся процессом, который происходит под давлением внешней среды и направлен на установление с ней динамического равновесия. В стволе мозга расположено четверохолмие, которое является местом обмена энергией моторного и сенсорного отделов нейронной сети. Стереотип движения глазного яблока формируется следующим образом. Предположим, сетчатка равномерно освещена и через нее прекращается поступление светового потока. Возникшее состояние равновесия сопровождается соответствующим тонусом глазодвигательных мышц, которые фиксируют глазное яблоко в определенном положении. Если в поле зрения появляется темная точка, то поток энергии, прошедший через сетчатку и бугорки четверохолмия, нарушает тонус мышц, заставляя глаз отклоняться в сторону. Через некоторое время сетчатка восстанавливает утраченное с внешней средой равновесие и блокирует прохождение энергии светового потока, поэтому глаз возвращается в исходное положение.
Текущие изменения освещенности порождают тремор (постоянные отклонения глазного яблока), который закрепляется в памяти бугорков четверохолмия, как форма поведения глазного яблока. Таким образом, в случае появления в нейронной сети избытка энергии, обусловленного внешними или внутренними (происходящими в поле сознания) процессами, происходит активация программ, записанных в памяти четверохолмия. Бугорки четверохолмия можно сравнить с клапаном, который отводит избыточную энергию из зрительного анализатора.
Если в поле зрения появляется объект более сложной формы, то его контур можно рассматривать как группу точек, расположенных по кольцу. В соответствии с логикой рассуждений, глаз “фиксирует” самую темную точку. После адаптации сетчатки глазное яблоко передвинется, уравновешивая соседнюю точку изображения и так далее. При достаточной скорости этого процесса, глаз начнет передвигаться по контуру предмета, нейтрализуя поступающий на его поверхность поток энергии, поэтому возникает возможность перескочить на менее яркий контур другого предмета, пробежав по которому он возвращается к исходному контуру и далее цикл повторяется. Таким образом, наряду с тремором появляется вторичная, более сложная форма поведения глаз. Еще более сложным стереотипом движения глаз является дрейф (“осмысленный” поиск объекта в поле зрения), появление которого обусловлено формированием филогенетической памяти орбитальной коры (особенности движения глаз, при рассматривании предметов, были описаны в 1957 году А.Я. Ярбусом).
По мере формирования равновесия между внешней средой и зрительным анализатором, наиболее эффективные движения (из общего набора хаотичных движений глазного яблока) кратно повторяются, что способствует их запоминанию. Поскольку набор потенциально возможных состояний равновесия с внешней средой консолидируется на мембране нейронов четверохолмия, движения глазного яблока обретают некоторую упорядоченность, определенный смысл. Программы движения глазного яблока, записанные в четверохолмии, являются одним из наиболее древних уровней развития ориентировочного рефлекса (еще более древним механизмом подобного рода является сброс энергии, через ядро продольного пучка, которое принимает участие в определении положения головы, как при зрительном, так и при слуховом воздействии). По нашему мнению, четверохолмие является примитивным аналогом коры мозга. Сформировавшийся в его памяти, по мере эволюции нейронной сети, стереотип движения глаз, является компонентом процесса мышления.
Благодаря тремору, поступление энергии через сетчатку в зрительный нерв становиться постоянным, вследствие чего в зрительном анализаторе формируется стабильная модель окружающего мира. Однако тремор глаз создает в зрительном анализаторе значительный избыток энергии, для нейтрализации которого необходимо дальнейшее усложнение его структуры (например, вертикальное или горизонтальное движение глаз способствует появлению на сетчатке вертикальных и горизонтальных линий, образуемых возбужденными фоторецепторами, что требует дополнительных механизмов для нейтрализации появившегося избытка энергии).
Как отмечалось выше, одним из вариантов нейтрализации избытка энергии в нервной системе является развитие новых слоев нейронной сети. Для каждого, вновь образовавшегося слоя нейронной сети, предыдущий слой, является внешней средой. С одной стороны это означает, что с усложнением нейронной сети расположенная в ней динамическая модель окружающей среды становиться точнее, а с другой стороны, нейроны очередного слоя нервной системы уравновешивают определенный параметр среды, то есть приобретают специализацию. Нейроны, способные реагировать на определенные параметры внешней среды, были открыты Д. Хьюбелом, при исследовании стриарной коры и названы нейронами-детекторами (131). По логике вещей, появление нейронов-детекторов должно происходить одновременно с формированием стереотипа движения глаз.
Для нейронов очередного слоя зрительного анализатора, события внешней среды, отражаемые в динамике одновременного возбуждения нейронов предыдущего слоя, представляют эту среду в упрощенном, отфильтрованном виде. Горизонтальное, вертикальное и прочие варианты движения глаз порождают, в соответствующих слоях рефлекторной сети, группу одновременно возбужденных фоторецепторов и уравновешивающих их нейронов. Каждый слой нейронной сети нейтрализует определенный узор информации предыдущего слоя, что выражается в образовании связи между группой одновременно возбужденных нейронов предыдущего слоя с одним нейроном последующего слоя.

Движение глазного яблока порождает в сетчатке группу одновременно возбужденных фоторецепторов, которые способствуют появлению, в вышерасположенном слое нейронной сети, нейрона реагирующих на эту группу. Таким образом, в зависимости от характера движения глаза, появляются нейроны, уравновешивающие различные сочетания активности фоторецепторов сетчатки.

При движении глаз, отрезок линии, совпадающий с направлением этого движения, способствует появлению в участке стриарной коры, условно обозначенном как второй слой, группы одновременно возбужденных нейронов. Ее появление формирует в третьем слое нейронной сети детектор направления.
Логика функционирования зрительного анализатора предполагает, что в результате движения глаз точечный источник света, во внешней среде, активирует группу линейно расположенных фоторецепторов, которые, в последующих слоях зрительного анализатора, активируют единичный нейрон (образ точки во внешней среде присутствует в нервной системе, несмотря на движение глаз). Линия, сочетание нескольких точек внешнего изображения, порождает нейрон-детектор направления. Для его появления необходимо, чтобы направление движения глаза совпало с пространственной ориентацией отрезка, что вызывает одновременную активацию нескольких “нейронов-точек”, которые уравновешивает один нейрон-детектор направления. Поскольку специализация нейронов-детекторов обусловлена тем, что глаз может двигаться только в одном направлении, каждое направление движения глазного яблока порождает свой класс уравновешивающих его нейронов. В конце концов, локальный участок сетчатки уравновешивается участком стриарной коры, который содержит нейроны-детекторы, нейтрализующие все возможные сочетания возбужденных нейронов (положения отрезка линии в поле зрения).

При движении глаз перпендикулярно отрезку в поле зрения деполяризуется участок рядом расположенных фоторецепторов, которые активируют горизонтальную клетку, в результате чего блокируются дальнейшее прохождение сигнала. Движение глаз вдоль отрезка не в состоянии активировать горизонтальную клетку и сетчатка остается проходимой.
Если, в результате движения глаз, перпендикулярного относительно внешнего расположения отрезка, на сетчатке появляется квадрат возбужденных фоторецепторов, изображение блокируется, так воспринимается, как фоновое освещение. Движением глаз можно объяснить парадокс, который привел в недоумение Д. Хьюбела. Он обратил внимание, что совмещение в диаскопе сетки вертикальных линий, предъявленных в один глаз, с сеткой горизонтальных линий, предъявленных в другой, воспринимается нами, не как ожидаемый по логике вещей образ решетки, а как хаотическое сочетание фрагментов горизонтальных и вертикальных линий. Объяснение простое, движение глаз (содружественное) исключает возможность одновременного перемещения глазного яблока в двух направлениях, следовательно, последовательность формирования образов в коре мозга исключает возможность их зрительного совмещения.

Нейроны-детекторы напрвления образуют в каждом учатке соответствующего слоя стриарной коры множество нейронов, способных уравновесить все радиальные положения отрезка линии в той или иной точке пристранства.

Сочетание одновременного возбуждения нейронов-детекторов направления определенного участка стриарной коры формирует нейроны-детекторы угла.
Если в поле зрения две линии (два радиуса) сходятся под углом (острым или тупым, как показано на схеме), то рядом расположенные детекторы линии возбуждаются одновременно, а в вышерасположенном слое нейронной сети появляются детекторы, реагирующие на угол или конец линии (если угол очень острый – формируется детектор "конца линии"). Таким образом, появляется новый класс простых детекторов, который уравновешивает избыток энергии появившийся в очередном слое стриарной коры.

Движение глазного яблока, совпадающее с направлением внешне расположенного отрезка, вызывает одновременное возбуждение однотипных нейронов направления нескольких “топографических ячеек” стриарной коры, что способствует появлению нейронов детекторов направления движения.
Если при движении стимула в одном и том же направлении (например, справа налево) несколько простых нейронов-детекторов последовательно возбуждается, то по мере увеличения длинны цепочки возбужденных нейронов, между ними могут возникнуть вставочные нейроны, вызывающие латеральное торможение. По мнению некоторых авторов, функциональная асимметрия системы вставочных нейронов, связанных с группой простых нейронов-детекторов направления, обуславливает возбуждение сложного нейрона-детектора направления движения, в ответ на определенное направление движения стимула. Схема, предложенная Х. Барлоу и У. Левиком, для объяснения детекторов направления движения, учитывает факт движения глаз в процессе поступления информации, в зрительный анализатор. В тоже время (с нашей точки зрения), основополагающим фактором появления этого вида детекторов является направление быстрого движения глаз, так при этом происходит практически одновременное возбуждение группы фоторецепторов и активация уравновешивающих их нейронов-детекторов на различных уровнях организации зрительного анализатора.

Схема, предложенная Х. Барлоу и У. Левиком, не совпадает с концепцией эволюции зрительного анализатора, как самоорганизующейся системы.
Чтобы уравновесить избыток энергии, поступающий в любой слой зрительного анализатора, должны сформироваться детекторы исчерпывающие все варианты своих “прародителей” во внешней среде. Следуя данной логике рассуждений, можно объяснить образование детекторов любых признаков внешней среды. В зрительном анализаторе существуют две параллельные системы off и on клеток, специализирующихся на светлых и темных изображениях. Для каждой системы справедливы вышеприведенные рассуждения. Чем больше нейронов детекторов участвует в установлении динамического равновесия со световым потоком, тем богаче зрительный образ. Процесс формирования образа сознания напоминает совмещение монохромных изображений при печати, для получения цветного изображения. Как видим, содержание сознания является динамической моделью окружающей среды, модулирующее влияние на которую оказывает прошлый опыт, наряду с активностью моторной части мозга, в свою очередь определяемой движением мышц и состоянием внутренней среды организма.
ПОСЛЕСЛОВИЕ
Уважаемый читатель, книга в Ваших руках, отвечает на многие вопросы, которые издавна волнуют человечество. Ознакомившись с нею, Вы поймете, как устроена материя, что такое сознание, мышление, память, эмоции, как устроен наш мозг. Автор, при изложении материала, использовал алгоритм рассуждений, который открывает пути к созданию универсальной системы философии. Несмотря на обширность затронутых проблем, уровень их изложения доступен не только аудитории, владеющей специализированными знаниями, но и рядовому читателю, склонному к философскому осмыслению действительности.