Мир будущего - В. А. Карачунский Философия движения - Биологическая эволюция. Развитие "живой материи"
1. Материя “живая” и “неживая”
Вопросы организации материи на биологическом уровне и ее отличие от неорганической природы, появились в глубокой древности и остаются актуальными до настоящего времени, так как до сих пор нет общепринятого мнения, по данному вопросу. Затронутая проблема касается обширного материала, который не укладываются в рамки одного научного направления и, тем не менее, применив к ней предложенные нами принципы организации материи, мы можем внести некоторую ясность по целому ряду вопросов. Одно из самых лаконичных определений органической материи принадлежит Ф.Энгельсу (142):
“Жизнь - это форма сосуществования белковых тел, существенным моментом которой является постоянный обмен с окружающей их внешней природой и которая прекращается вместе с прекращением этого обмена веществ, ведя за собой разложение белка”.
Несмотря на множество определений феномена “жизнь”, которые появились в последствии, ни одно из них по лаконичности, образности и как увидим дальше, скрытому эволюционному смыслу, не сравнимо с таковым у Ф. Энгельса. В философии широко используется подход, согласно которому жизнь следует рассматривать, как высшую форму существования материи. В тоже время, с точки зрения диалектики, оно не совсем корректно, так как напрашивается вывод, что дальше материи развиваться некуда.
В работе “Происхождение видов”, Ч.Дарвин сформулировал основные законы, общие для различных форм проявления жизни. По его мнению, к ним относятся рост, воспроизведение, наследственность и изменчивость. Однако эти законы применимы к эволюционным процессам не только в живой, но и в неживой природе. Геофизикам хорошо известно, что в определенных условиях планеты, способны зарождаться из вещества вселенной, развивается и усложняется, поэтому некоторые специалисты сравнивают планету с живым организмом. Многие неорганические образования на Земле исчезают и появляются вновь. С другой стороны, находящийся вне тканей организма хозяина вирус, не является обладателем каких либо атрибутов живого организма (в вирусной частице отсутствуют метаболические процессы, она не способна размножаться и так далее).
С точки зрения диалектического материализма, появление жизни на земле, является результатом усложнения вещества Земли. Позицию, согласно которой, для живой и неживой материи характерны общие принципы строения и развития, занимали многие естествоиспытатели. Например, в восемнадцатом веке, английский врач Д. Гартли сделал предположение, что в нервной системе существуют циклические процессы, которые подобно процессам движения планет солнечной системы по замкнутым траекториям, обеспечивают ее стабильность (47). Австрийский астрофизик Э.Шредингер утверждал, что живые организмы, как бы питаются отрицательной энтропией, извлекая ее, из окружающей среды и увеличивая возрастание положительной энтропии (2). Все эти идеи, высказанные в свое время специалистами различного профиля, предполагают рассмотрение жизни, как части динамических процессов происходящих во вселенной, но общей теории из этого не следует.
Известный биохимик В.А. Энгельгардт, считает, что основными свойствами живой системы являются ее способность к воспроизведению и изменчивости. В качестве аргументации он использует данные, полученные в процессе исследования генетического материала клетки. Запись наследственных свойств, кодирование признаков организма, необходимое для воспроизведения, осуществляется с помощью ДНК и РНК. В самом процессе репродукции, непременно принимают участие белки-ферменты, а сам процесс осуществляется в очень специфической среде, цитоплазме. Если мы будем следовать этой логике, то можем прийти к выводу, что “живыми” являются не отдельные молекулы ДНК, белка или РНК, а система, в которой они объединяются в нечто целое. Именно ей, данной системе, будут свойственны воспроизведение и изменчивость.
Несмотря на то, что науке хорошо известно из чего состоят живые организмы, как организованы процессы, обеспечивающие их устойчивость в окружающей среде, наука не может установить тот Рубикон, который отделяет “живое” от “неживого”. Предлагаемая нами теория движения, в какой то мере развивает определение понятия жизнь, предложенное Ф. Энгельсом, рассматривая ее, как очередной уровень организации неживой природы, формируемый циклическим движением нуклеотидов и аминокислот.
2. Место органического вещества в общей динамике развития материи
Одна из наиболее известных гипотез о происхождения жизни не Земле, изложена советским биохимиком А.И.Опариным (монография “Происхождение жизни”, опубликованная в 1924 году). Суть этой гипотезы, соответствует тенденциям материалистического подхода и заключается в предположении, что простейшие организмы появилась на Земле в результате длительного процесса химической эволюции и последующего действия естественного отбора на уровне многомолекулярных образований, предклеточных структур.
Вторая основополагающая гипотеза, направленная на разгадку феномена жизни, связана с работами английского физика Ф. Крика, специалиста по вопросам молекулярной биологии и посвящена рассмотрению принципов организации генетического материала. В 1953 году Крик, совместно с Дж. Уотсоном, описал устройство молекулы дезоксирибонуклеиновой кислоты, из которого становилось понятно, чем обусловлена ее избыточная устойчивость в среде прочих молекулярных образований. Согласно этим представлениям, молекула ДНК являет собой двойную спираль, построенную из комплементарных друг другу цепей дезоксирибонуклеотидов. В процессе репликации (удвоения), водородные связи между парами нуклеотидов разрываются и к ним присоединяются дезоксинуклеозидтрифосфаты, присутствующие в окружающей среде и комплементарные соответствующим дезоксирибонуклеотидам (процесс воспроизведения). Следуя вышеприведенным гипотезам, приходим к выводу:
Условием возникновения жизни является появление особых, избыточно устойчивых в окружающей среде форм химической организации материи
Как показано в первой части книги, процессу эволюции материи свойствен как прогресс, так и регресс, который проявляется в образовании или разрушении уровней ее организации. Для процесса усложнения материи характерно, что в него включаются в неизменном виде элементы предыдущего. Например, солнце являет собой плазменный шар, в котором устойчивыми элементами являются ионы. В результате эволюции небольшого количества вещества, оторвавшегося от солнца, образовалась наша планета, в которой ионы плазмы вошли в состав более сложных структур, атомов, которые составляют основную массу вещества нашей планеты.
В процессе образования уровней движения материи, просматриваются общие закономерности. На очередном, вновь образованном уровне движения, появляется большее разнообразие циклических форм, чем на предыдущем. В тоже время, вновь образованные циклы движения требуют более “рафинированных” условий существования, в сравнении со своими предшественниками. Например, циклы движения биологического уровня не способны существовать в тех условиях, в которых существовали циклические элементы предыдущего уровня (избыток энергии присутствующий на химическом уровне организации вещества, при формировании биологического уровня, частично переходит в потенциальное состояние, способствуя усложнению материи, поэтому вновь появившиеся элементы движения биологического уровня, обладают меньшей скоростью перемещения и следовательно имеют более низкий порог устойчивости к избытку энергии в окружающей среде).
Таким образом, материя представляет собой многоуровневую динамическую систему, каждый “этаж” которой образован одному ему присущими формами циклического перемещения элементов предыдущего уровня (характер движения элементов определяет субъективное восприятие материи на данном уровне). Циклы движения образующие вещество на молекулярном уровне содержат атомы, которые становятся неуничтожаемыми элементами, появившимися на предыдущем уровне. Молекулярный уровень организации материи можно условно обозначить следующим образом:

Молекула, элементарная структура очередного уровня организации материи, с точки зрения циклической формы движения.
Следует подчеркнуть, что устойчивым, на том или ином уровне движения, является не участок пространства, а присущий данному участку характер циклического движения находящихся в нем элементов. Например, неуничтожаемым элементом молекулярного уровня движения является не конкретная молекула воды, а характер движения атомов, из которых она состоит (данную молекулу воды можно разрушить, но пока существует молекулярный уровень движения, вода, как устойчивая циклическая форма движения, будет присутствовать в общей массе движения).
На определенном этапе формирования устойчивых циклов движения, состоящих из элементов молекулярного уровня, произошло появление дезоксирибонуклеиновой кислоты, неуничтожаемого элемента биологического уровня. Дезоксирибонуклеиновая кислота состоит из двух нитей, в формировании каждой из которых принимают участие четыре азотистых основания – аденин, тимин, гуанин, цитозин. Последовательность их соединения является генетическим кодом организма. Нуклеотиды, так же как и сложные молекулярные образования на их основе, принадлежат молекулярному уровню организации движения. Появление биологического уровня знаменуется парным объединением спирально закрученных нитей дезоксирибонуклеиновой кислоты, в которых азотистые основания расположены таким образом, что напротив аденина, может находиться только тимин, а против гуанина, только цитозин. Подобная комплементарность нуклеотидов приводит к тому, что последовательность их расположения в одной цепи, однозначно определяет их последовательность в другой.

Две цепочки, состоящие из азотистых оснований, образуют между собой структуру, более устойчивую в окружающей среде, чем каждая из нитей ДНК в отдельности, которая становиться устойчивым элементом биологического уровня движения.
Вновь образовавшаяся конструкция из сдвоенных нитей ДНК, способна восстанавливать потерянный фрагмент одной из нитей, захватывая его из окружающей среды, поэтому, при прочих равных условиях, она более устойчива во внешней среде, чем родственные ей образования.
Как видим, органическая материя является следствием развития неорганической материи, в которой появляются неуничтожаемые элементы, из которых начинает формироваться очередной, биологический уровень движения. По мере усложнения материи в ней появляются тенденции, согласно которым процесс эволюции биологической формы движения замедляется, а процесс эволюции окружающей среды наоборот, ускоряется (человек, адаптируя к своим потребностям окружающую среду, теряет, в определенной мере, необходимость к ней приспосабливаться).
Первым уровнем биологической формы движения следует считать появление сдвоенной цепочки нуклеотидов в аминокислотной среде. По мере развития органической материи в ней формируются очередные уровни движения, имеющие свои неуничтожаемые элементы, клетка, многоклеточный организм, социум (для каждой устойчивой формы движения характерна своя логика перемещения составляющих ее элементов, позволяющая относить ее к тому или иному классу). Следовательно:
Появление жизни являет собой закономерный этап усложнения динамической системы материи до уровня, на котором появилась ДНК, устойчивый элемент химического уровня, образованный азотистыми основаниями, способный формировать новый, биологический уровень движения
3. Циклическая форма движения в органическом веществе
В самом начале эволюции живой природы, согласно теории А.И. Опарина, нуклеотиды плавали в аминокислотной среде, где устойчивые варианты дезоксирибонуклеиновой кислоты были окружены “обломками” менее стойких в среде “собратьев” (нуклеотидами и аминокислотами). Парное соединение спиралей дезоксирибонуклеиновой кислоты представляет конструкцию, более устойчивую во внешней среде, чем окружающие ее молекулярные образования. При существовании этой конструкции в родственной ей аминокислотной среде, с ней взаимодействуют белковые образования (цепочки аминокислот). Согласно логике движения материи, две взаимодействующие циклические системы стремятся к равновесию, идеальным исходом которого будет их полная идентичность (локальная энтропия). Дезоксирибонуклеиновая кислота и аминокислоты близки по химическому составу, и будучи изолированы от окружающей среды, вероятно, образовали бы однородную массу. В реальных условиях, внешняя среда постоянно нарушает равновесие между дезоксирибонуклеиновой кислотой и окружающими ее молекулами аминокислот.

Дезоксирибонуклеиновая кислота, стремясь к равновесию с окружающей средой и обладая повышенной к ней устойчивостью, сама не меняется, а заставляет объединяться между собой аминокислоты, формируя белковые тела.
Фактически, дезоксирибонуклеиновая кислота, взаимодействуя с аминокислотами, образует вокруг себя защитную белковую оболочку, непосредственно контактирующую с внешней средой (белки превращаются в посредников, между дезоксирибонуклеиновой кислотой и внешней средой).

Белковые тела образуют вокруг молекулы дезоксирибонуклеиновой кислоты циклически организованную форму движения, непосредственно контактирующую с внешней средой.
Взаимодействующие между собой дезоксирибонуклеиновая кислота и белковые тела соизмеримы по массе и составу, поэтому, стремясь к равновесию, белковые тела повторяют логику движения дезоксирибонуклеиновой кислоты, превращаясь в ее приблизительную копию. Это позволяет, схематично обозначить дезоксирибонуклеиновую кислоту и белковые образования, как ДНК 1 и ДНК 2.
Следовательно, живая материя, это многослойная структура, в центре которой находится ДНК. В процессе взаимодействия с окружающей средой она обретает упорядоченность, образуя оболочку, которая становиться посредником между внешней средой о ДНК. Каждый, сформировавшийся слой, является приблизительной копией ДНК (чем ближе слой расположен к самой ДНК, тем эта копия точнее).

Две нити дезоксирибонуклеиновой кислоты формируют, из окружающего их белкового материала, свою приблизительную копию. Образовавшаяся оболочка становиться посредником между генетическим материалом и внешней средой.
Подобное представление об организации живых организмов помогает определить логику их развития, функционирования и строения. Оболочка и цитоплазма клетки, контактирующие с внешней средой, являются приблизительной копией хромосом (комплекса, образованного дезоксирибонуклеиновыми кислотами). Цитоплазма, внутренняя часть белковой системы клетки, непосредственно контактирует с генетическим материалом и является более точной его копией. Оболочка менее похожа на генетический материал, чем цитоплазма, так как контактирует с внешней средой непосредственно и, соответственно более удалена от генетического материала (по этой причине, клетки тканей организма отличаются друг от друга в большей степени по структуре оболочки, чем по строению цитоплазмы).
В действительности, вышеописанный принцип организации динамических процессов в клетке, имеет более сложную, многоуровневую структуру. Парные нити дезоксирибонуклеиновой кислоты, объединяются между собой, образуя гигантскую молекулу, хромосому. Хромосомы в клетке, в свою очередь, образуют парные структуры. Весь генетический материал формирует единую динамическую структуру, ядро клетки. Таким образом, нити ДНК дублируются несколько раз, что еще больше увеличивает их устойчивость к внешним воздействиям. Эта устойчивость проявляется в растущей способности генетического материала активно изменять среду вокруг себя. Белки и рибонуклеиновая кислота, образуют вокруг хромосом динамическую оболочку, которая уравновешивает внешние воздействия. Хромосома закручена в спираль. Там где этот процесс выражен сильнее, контакт с окружающей средой минимален. В местах, где спираль распрямляется, ее участки получают возможность активно взаимодействовать с внешней средой. Сочетание активных и неактивных участков хромосомы уравновешивает дежурное состояние среды вокруг клетки.
Наиболее динамично, под воздействием окружающей среды, изменяется оболочка клетки. Перестройка структуры цитоплазмы происходят значительно медленнее. Еще более медленным процессом, является эволюция хромосом и дезоксирибонуклеиновых кислот. Затихающие “волны эволюции”, которые достигают центра клетки, воздействуя на структуру одной из нитей дезоксирибонуклеиновой кислоты, встречают противодействие со стороны другой нити, которая стремится восстановить структуру своей соседки. Таким образом, каждый вновь образованный уровень движения, является оболочкой для предыдущего и охраняет его от внешнего воздействия, что объясняет способность живых организмов к самовосстановлению. С этой точки зрения, “регенерация”, это динамический эквивалент понятия “устойчивость” к внешнему воздействию, для многослойной системы, что заложено в динамических особенностях материи. Особо выражена эта способность у организмов стоящих на более низкой ступени развития и имеющих меньшее количество уровней движения в своей структуре.
По мере усложнения структуры слоев-посредников и самого генетического материала, нарастает устойчивость биологической системы, к изменениям окружающей среды и все продолжительнее становятся периоды динамического равновесия. Так, клетки многоклеточного организма реагируют на внешние воздействия не изменением своей структуры, а изменением активности тех или иных участков хромосомы и соответствующей функциональной перестройкой структур клетки, то есть имеют “запас прочности” в потенциальном виде. Поэтому, чем сложнее структура живого организма, тем медленнее эволюционирует его генетический материал и тем больше в его внешней оболочке появляется потенциальных возможностей уравновешивать внешнюю среду. В конечном счете, можно утверждать, что стремление к равновесию, самовосстановление память, являются различными точками зрения на один и тот же процесс (память, это изменения в структуре данного уровня движения, которые не принимают непосредственного участия в становлении равновесия с текущим состоянием внешней среды).
4. Схема эволюции живой материи
Одним из наиболее простых вариантов циклического движения биологического уровня, в основе которого лежит элементарная частица - дезоксирибонуклеиновая кислота, является взаимодействие этой частицы с белковыми молекулами, которые образуют вокруг нее оболочку, непосредственно контактирующую с внешней средой. Необходимо уточнить, что элементарная частица, это не нечто-то неподвижное, а циклическое движение определенного характера, которое сохраняется в генетическом материале любой сложности (элементарная форма циклического движения азотистых образований, по мере образования новых уровней циклического движения, превращается в комплекс хромосом).

Парная нить дезоксирибонуклеиновых кислот образует с белками динамическую систему, более устойчивую во внешней среде, чем родственные ей молекулы.
Результатом взаимодействия двух систем, является стремление к установлению между ними динамического и структурного равенства, поэтому генетический материал обретает логику движения внешней среды, а внешняя среда, обретает логику движения генетического материала. Более высокий уровень устойчивости парных молекул дезоксирибонуклеиновой кислоты, проявляется в способности усложнять окружающую их внешнюю среду и способствует появлению в ней структур, имеющих характер движения генетического материала.

Оболочка вируса повторяет логику движения его генетического материала, поэтому обе системы могут быть обозначены как ДНК 1 и ДНК 2.
Примером начальной стадии развития живой материи является вирус. Вирусная частица состоит из нити дезоксирибонуклеиновой кислоты и окружающей ее оболочки, состоящей из белков (каспид). С точки зрения своей динамической организации, оболочка вируса повторяет логику движения, присущую его генетическому материалу (оболочку можно сравнить с системой рычагов, объединяющих в единой системе логику движения внешней среды и молекулы дезоксирибонуклеиновой кислоты).
Следующим, условно выделенным уровнем организации биологической формы движения, является клетка.

В структуре клетки можно выделить несколько уровней организации, имеющих общую логику движения, ДНК 1, ДНК 2 и ДНК 3.
Генетический материал клетки формирует вокруг себя оболочку из белковых тел и рибонуклеиновых кислот (образуется белковая система клетки, являющаяся посредником между текущими изменениями среды и генетическим материалом). В конечном счете, динамика процессов в белковой системе клетки частично копирует динамику процессов в ее генетическом материале. Первой “копией” генетического материала становится кариоплазма и оболочка ядра клетки, второй, менее точной его “копией”, становиться цитоплазма и оболочка клетки.
Попробуем проследить данную логику рассуждений, применительно к многоклеточному организму. Многоклеточный организм может быть условно представлен в виде сферы, стенка которой, прототип тканей организма, образована клетками. Внутри такой сферы расположена полость, которая будет средой общей для всех ее клеток (в отличие о среды вокруг сферы, ее внутренняя среда имеет конечные размеры). Следовательно, полость в внутри сферы можно обозначить, как внутреннюю внешнюю среду (ВВС) . Тогда среду вокруг сферы следует определить, как наружную внешнюю среду (НВС) . Если динамическую структуру, образованную генетическим материалом клетки, обозначить как ДНК 1 , то ее клеточную оболочку можно обозначить, как ДНК 2, а логику процессов во внутренней среде организма, как ДНК 3 .

Многоклеточный организм может быть представлен в виде скопления клеток, внутри которого сформировалась замкнутая полость, внутренняя внешняя среда (ВВС). Характер процессов внутри этой полости отражает логику динамики генетического материала организма и может быть обозначен, как ДНК 3.
Масса клеток организма соизмерима с массой внутренней среды организма, так как последняя имеет конечные размеры. По этой причине внутренней среде организма передаются общие черты логики движения его клеток, которая в своей основе имеет логику движения генетического материала. Становится понятней, что такое иммунитет. Кровь и тканевая жидкость, являются средой, общей для всех клеток организма и несут в себе отпечаток его генетического материала. В свою очередь все белковые образования данного организма содержат динамические закономерности генотипа, содержащегося в его клетках. Если во внутреннюю среду (кровь, тканевую жидкость) попадает чужеродный белок, объект с нехарактерной для генотипа данного организма динамической структурой, он начинает деформировать существующий во внутренней среде организма характер движения. В свою очередь, внутренняя среда пытается воспрепятствовать этому процессу и деформирует логику движения чужеродного генетического материала. В конечном счете, побеждает объект с большей массой, то есть внутренняя среда организма, что приводит к разрушению чужеродного белка.

На схеме показано, что клетка организма имеет три варианта взаимодействия с окружающими ее структурами. Участок А контактирует с наружной внешней средой, участок В с оболочкой соседней клетки, а участок С соприкасается с внутренней внешней средой.
При рассмотрении взаимоотношений среднестатистической клетки организма с окружающими ее образованиями, можем установить определенные закономерности. Оболочка клетки многоклеточного организма, в участках В и С, формируется под воздействием генетического материала и отражает его логику. Воздействие на клетку в участке А будет зависеть от особенностей параметров среды вокруг организма. Таким образом, эволюция отдельных клеток организма определяется воздействием на них наружной внешней среды и противодействием со стороны генетического материала, стабилизирующего динамику клеточных процессов. Следовательно, ткани организма являются очередной оболочкой, которая формируется вокруг дезоксирибонуклеиновой кислоты.
На уровне одноклеточного организма определяется функциональная разобщенность отдельных участков мембраны. Одни из них служат для входа энергии и вещества из клетки, другие наоборот, способствуют их поглощению. Специализация поверхности мембраны зависит от особенностей пространственного расположения клетки, а также от последовательности и кратности внешних воздействий. В многоклеточном организме, роль участков поглощения и обратного возвращения энергии внешней среды принадлежит многоклеточным образованиям, так как для эффективности этого процесса в масштабах целого организма необходимо синхронная работа нескольких клеток. Поглощение энергии и образование вещества (новых уровней движения), происходит во внутренней среде организма, своеобразном клеточном буфере, возможности которого значительно превосходят потенциал единичной клетки. Этот процесс, субъективно воспринимаемый, как обмен веществом и энергией между организмом и средой, является проявлением совместной, циклически организованной системы движения данного участка пространства.
Поскольку, любая, циклически организованная, многоуровневая система является частью целого, в ней периодически появляется избыток энергии, который оседает в структурах организма, заставляя их эволюционировать. Каждый, вновь образовавшийся уровень биологической системы, берет на себя часть внешней энергии, замедляя эволюцию более низких ее уровней.
Появление нервной системы, приводит к образованию очередной оболочки, уровня движения, представленного циклически организованными потоками импульсов на мембранах нейронов, на которые, через сенсорные анализаторы, поступают волны избытка энергии из среды, окружающей организм. Основная масса энергии, которая поступила в нервную систему, выводиться в окружающую среду, в виде хаотической активности мышечной ткани и только небольшая ее часть расходуется на эволюцию нейронной сети. Фактически, нервная система является устройством, которое преобразует энергию внешней среды в энергию поведенческой активности.
Появление нервной системы происходит под воздействием окружающей среды, в результате эволюции эпителиальной ткани (46). Оболочка клеток эпителия, вблизи мышечных клеток, приобретает свойства проводимости (энергия, поступающая на оболочку клетки, находит универсальный выход в мышечной ткани, поэтому появление и развитие нервной и мышечной ткани происходит параллельно). В местах контакта с внешней средой, оболочка клетки превращается в примитивный фильтр, особенности которого определяются характером и кратностью внешнего воздействия. Участок оболочки клетки, контактирующий с окружающей средой, эволюционируют быстрее, оставшейся ее части и приобретает специализацию по противодействию определенному виду избытка энергии, характерного для данной точки пространства. Со временем, специализированные участки клеток превращаются в рецепторы, реагирующие на химическое, механическое, звуковое световое и прочие виды внешнего воздействия.
Таким образом, оболочка эпителиальной клетки обретает свойства проводимости, а сама клетка превращается в нейрон. Отдельные нейроны объединяются между собой в нейронную сеть, внутри которой формируются синапсы, наиболее распространенные образования в нервной системе (по сути, синапс, является рецептором, воспринимающим энергию от мембраны соседнего нейрона). Синапс образован участком мембраны нейрона, отдающего энергию (пресинаптическая мембрана) и участком мембраны нейрона ее принимающего (постсинаптическая мембрана). Среда между мембранами, это своеобразный “морской залив”, имеющий свой “микроклимат” и не теряющий связь с “открытым морем”, внутренней средой организма.
Избыток энергии внешней среды, в масштабе многоклеточного организма, поступает в нервную систему, через группу однотипных рецепторов (рецепторное поле). Количество поступающей энергии определяется способностью мышечной ткани отдавать ее в окружающую среду, поэтому развитие структуры нервной и мышечной тканей происходит в определенном соответствии друг другу.

Нервная и мышечная система, это функциональное образование в тканях организма, пропускающее через организм волны избытка энергии, возникающие во внешней среде. Принцип организации данного образования находит свое выражение в схеме рефлекторной дуги.
Мир вокруг нас находиться в динамическом равновесии (образует циклическую форму движения). Даже в тех случаях, когда отдельные фрагменты перемещения объектов мы относим к линейному движению, факт того, что мы его воспринимаем, заставляет думать о циклическом характере наблюдаемого процесса, в системе более высокого порядка. Например, хождение по магазинам, прогулки на природе, в нашем восприятии процесс линейный. В тоже время, вне зависимости от нашего маршрута, мы, рано или поздно, должны будем вернуться обратно. Следовательно, любое наше перемещение в этом мире, есть процесс циклический. Движение мышц, классический пример циклического процесса. Выход энергии из организма не может быть обеспечен линейным механизмом, так как после сокращения мышца не способна выводить энергию и должна обрести изначальное состояние. Одна мышечная единица обеспечивает фрагментарное выведение энергии из организма. По мере развития мышечной системы, в ней образуются участки, находящиеся в антагонистичных отношениях, что обеспечивает возможность непрерывного выхода энергии (происходит сброс энергии в мышцу, которая находится в состоянии расслабления до тех пор, пока дальнейшее ее поступление становится невозможным и энергия начинает поступать в мышцу антагонист). Появление мышц антагонистов способствует усложнению поведения организма и формированию очередного уровня его организации.

Фрагментарное поступление и выход энергии в отдельные мышцы, приводит к колебанию в них ее уровня. Сочетание активности мышц, находящихся в противоположных энергетических состояниях, обеспечивает непрерывный выход энергии во внешнюю среду.
Таким образом, в нервной ткани образуется оболочка на пути воздействия внешней среды, состоящая из циклических потоков импульсов на мембране нейронов.

В нервной системе образуется динамическая модель тканей организма, которая образует единую систему с “виртуальной” копией внешней среды (наружной и внутренней).
Потоки импульсов, под воздействием поступающего в них избытка энергии, способствует появлению в нервной системе динамической копии контактирующих с ней систем: окружающей среды, внутренней среды организма и его тканей (две взаимодействующие циклические формы движения неизбежно копируют друг друга). В тоже время, копия внешней среды в тканях организма согласуется не только с воздействиями внешней среды, но и с особенностями выхода энергии из нервной системы (поведенческим континуумом). Таким образом, сознание (копия внешней среды), характеристика процесса, ориентированного на активность организма.
Избыток энергии, поступающий в нервную систему, неизбежно оказывает влияние на функциональное и анатомическое состояние мембраны нейронов. Изменения структуры нервной системы, за длительный период времени, создают в нейронной сети образования, которые уравновешивают не только текущие, но и вероятностные состояния среды, что позволяет говорить о феномене памяти.

В основании биологического уровня находится генетический материал, который, взаимодействуя с внешней средой, формирует множество оболочек, сообщая им логику своего движения.
На начальных стадиях формирования нервной системы, волны избытка энергии, преобразованные в хаотическую мышечную активность, влияют на внешнюю среду не более чем световой поток на поверхность зеркала, от которого он отражается. По формирования памяти, изменений структуры нервной системы, отражающих периоды временного равновесия с окружающей средой, активность организма становиться все более упорядоченной. Значительное увеличение памяти приводит к усложнению поведения, которое начинает активно изменять среду, окружающую организм. Таким образом, поведенческий континуум образует вокруг организма оболочку, уравновешивающую избыток энергии во внешней среде. Данные рассуждения позволяют прийти к определенным выводам:
Живая материя – это закономерный уровень организации неорганического вещества. Он знаменуется появлением сдвоенной молекулы дезоксирибонуклеиновой кислоты, которая избыточно устойчива в среде родственных ей молекулярных образований и начинает их “организовывать”, формируя вокруг себя уровни движения, уравновешивающие воздействие внешней среды. Появление нервно-мышечной системы формирует вокруг организма “поведенческую” оболочку, которая уравновешивает определенную часть избытка внешней энергии.
Представление о мутациях, как случайном факторе эволюции живых организмов, иллюзия (идея мутаций применима к двуполому размножению). Внешняя среда оказывает непрерывное давление на генетический материал организма, через многочисленные, окружающие его оболочки. Если величина избытка энергии, непрерывно поступающего в биологическую систему, превышает определенный уровень, происходит модификация оболочки. Генетический материал, защищенный от воздействия внешней среды многочисленными оболочками, наиболее “уязвим” для внешних воздействий в половых клетках, где гаплоидный набор хромосом соприкасается с внутренней средой организма, которая, в какой-то мере является его динамической копией. Таким образом, внутренняя среда организма передает на генетический материал, в половых клетках, волны избытка энергии от отдельных участков организма, которые в свою очередь подвергаются избыточному давлению со стороны внешней среды.
Энергия внешней среды, организуя циклические потоки импульсов на мембранах нейронов, способствуют появлению в нервной системе динамической модели, которая уравновешивает все контактирующие с ней системы. Изменения внешней среды требуют периодического обновления структуры потоков импульсов, что приводит к перестройке поверхности мембраны нейронов, которая начинает уравновешивать состояния внешней среды на протяжении все большего отрезка времени, что лежит в основе феномена памяти.
С точки зрения внешнего наблюдателя, образование в динамической системе нового уровня движения, при сохранности ранее существовавших уровней, может рассматриваться, как активность системы. Таким образом, не только биологическая, но и любая устойчивая система, обладает некоторой активностью по отношении к окружающей ее среде.
5. Закономерности поступления информации в нервную систему
Используя общепринятые подходы трудно объяснить отдельные феномены строения и функционирования мозга. Один из теоретических парадоксов, возникших в связи с изучением нервной системы, рассматривается в статье П.К. Анохина, посвященной интегративной деятельности нейрона (7). Суть проблемы заключается в следующем. Нейрон, состоит из сомы, дендритов и аксона. На его поверхности сходится в среднем до 5000 синаптических окончаний от соседних клеток. Сложный узор возбужденных, заторможенных и интактных синапсов, должен быть проанализирован мембраной и преобразован в потенциал действия, направляемый через аксон к другим нейронам. П.К.Анохин совершенно справедливо считает, что для интеграции поступившей на поверхность мембраны информации требуются дополнительные механизмы, не связанные с особенностями ее строения. Как выход из создавшегося положения он предполагает, что интеграция информации поступающей на мембрану нейрона обеспечивается не ее структурами, а сложной химией протоплазмы нейрона и генетического материала, с которой она связана. Внутриклеточный путь проведения информации содержит субсинаптическую мембрану, субсинаптические цистерны, микротрубочки дендрита, аксонный холмик. При прохождении через центр нейрона информация модулируется химическими процессами с участием ядерной РНК.
По нашему мнению, предположение о том, что генетический материал одного нейрона способен “разобраться” и оказать влияние на организацию процессов всей нервной системы, принципиально неверно. Условия существования клеток в нервной системе приблизительно одинаковы. Их мембрана уравновешивает находящийся рядом избыток энергии и не нуждается для этого в дополнительной информации о состоянии всей нервной системы. Еще меньше в ней нуждается генетический материал нейрона, тем более что он получает ее в отфильтрованном поверхностью мембраны, упрощенном виде.
Поле сознания, состоящее из кольцевых потоков импульсов, стремящихся к равновесию с окружающими средами, повторяет логику их движения. Таким образом, в нем образуются динамические копии внешней, внутренней среды и тканей организма (в каждой песчинке отражается вся вселенная и чем сложнее песчинка устроена, тем копия ближе к оригиналу). Гипотеза П.К. Анохина предполагает непосредственное воздействие генетического материала нейрона на поле сознания. Согласно нашим представлениям, посредником между генетическим материалом организма и полем сознания является цитоплазма клеток и внутренняя среда организма, увеличенная копия его генома (представительство внутренней среды взаимодействует с копией внешней среды в поле сознания).
Взаимосвязь генов и внешней среды осуществляется через целый ряд посредников: цитоплазму клеток, внутреннюю среду организма, нервную систему, циклические потоки импульсов в нейронной сети. Все перечисленные оболочки по-своему организуют свои взаимоотношения с внешней средой, уравновешивая часть ее воздействий. Оболочка, представленная кольцевыми потоками импульсов на суммарной мембране нейронов, образует устойчивую динамическую структуру, поле сознания, которая контактирует непосредственно с внешней средой. Оболочка поля сознания отражает волны избытка энергии, преобразуя их в мышечную активность, которая вначале является хаотической, но со временем обретает порядок (стереотип поведения). Энергия, поступившая в поле сознания, деформирует его. В свою очередь, это нарушает существующие взаимоотношения сознания с мембраной нейронов, которая, восстанавливая утраченное равновесие, меняет свою структуру (феномен памяти).
При изучении особенностей циркуляции импульсов, в условно выбранном участке нервной системы, мы часто не учитываем, что они являются частью целого, динамика которого отлична от динамики его частей (мы воспринимаем изображение на экране телевизора целиком, а не как сумму отдельных точек различной яркости). Соприкасаясь с различными средами, система потоков импульсов, составляющих поле сознания, копирует логику их движения, объединяя между собой. Вектор движения потоков импульсов в нервной системе определяется, с одной стороны, особенностями избыточной энергии в нее поступающей, а с другой, возможностью ее выхода посредством мышечной активности. Таким образом, поле сознания это оболочка, где устойчивые циклические формы движения, образы сознания и программы поведения, передвигаясь в рамках единой системы, уравновешивают воздействия внешней среды.
Существующие методики исследования нервной системы, как системы состоящей из многих нейронов, ориентированы на изучение одного или нескольких нейронов, а не мозга в целом (одновременное исследование активности большой группы клеток мозга затруднительно, с технической точки зрения). Следствием такого подхода является традиция, согласно которой функциональной единицей нервной системы считается нейрон, а не нервный узел. В контексте нашего изложения, предпочтительней рассматривать структуру нервной системы в виде объединения нервных узлов. Нервный узел, это, прежде всего, функциональная единица нервной системы, которую нельзя рассматривать с чисто анатомической точки зрения, как сумму клеток. Одновременность поступления энергии одного вида в рецепторное поле и последующее ее продвижение по многослойной структуре нейронной сети, обеспечивается цепочкой родственных нервных узлов, индивидуальные особенности которых определяются возможностью выхода поступившей в них энергии. Таким образом, при выделении нервного узла, из общей массы нейронов, на первый план следует поставить связь с группой периферических рецепторов одного вида.
Благодаря однородности составляющих его элементов, нервный узел реагирует на изменения параметров окружающей среды как нечто целое (при изменении обстановки вокруг организма нарушается равновесие между рецепторами и определенными физическими параметрами, что изменяет активность всех нейронов данного узла). Таким образом, для характеристики функционального состояния нервного узла, можно ввести понятие узор информации.
Узор информации, это сочетание уровня пульсации нейронов данного нервного узла, уравновешивающее определенное состояние внешней среды
Узор информации нервного узла можно сравнить с таковым в калейдоскопе. Пока на калейдоскоп не оказывается внешнее воздействие, видимый в нем узор будет устойчив. При вращении калейдоскопа, на месте исходного образуется новый узор. То же происходит и в нервном узле, где сочетание параметров внешней среды определяет уровень возбуждения отдельных нейронов узла (качество узора информации), а порог возбудимости отдельных рецепторов определяет толерантность узора к изменениям окружающей обстановки (устойчивость узора информации).
Представление о том, что нервная система получает информацию от внешней среды и использует ее в своих целях, направляет ход наших мыслей по ложному пути. Сразу напрашивается ассоциация, что в нервной системе существуют структуры, для которых эта информация предназначается. Нервная система, это не проводник энергии внешней среды, а оболочка вокруг генетического материала организма, которая уравновешивает часть внешних воздействий. Например, П.К. Анохин недоумевает, зачем нейрону необходима такая сложная система передачи информации, включающая в себя сложнейшие химические преобразования на всем пути следования импульса (7). Если рассматривать нейрон, как передаточную станцию для информации, которая “кем-то” ожидается в центре нервной системы, это недоумение вполне понятно. С нашей точки зрения, все можно объяснить проще. Многоплановость структуры синаптического аппарата между нейронами, вполне оправдана, если не относиться к ней, как к проводнику внешнего воздействия, а как к барьеру на пути этого воздействия. Чем значительней воздействие, тем сложнее устроен барьер его отражающий (синапс, это структура, при прохождении через которую расходуется энергия внешнего воздействия и чем значительный поток внешней энергии, тем сложнее структура синапса).
6. Появление клеток нервной и мышечной ткани
Попробуем проследить логику формирования нервно-мышечной системы. Первым шагом на этом пути будет понимание необходимости замены принципа проводимости, на принцип противодействия. Нервно-мышечная система, это оболочка, которая встречает внешние воздействия и уравновешивает их, меняя свою структуру. Проводимость необходимо оценивать не в изолированном виде, а как первую фазу поступления энергии от внешней среды. Вторая фаза этого процесса представлена явлениями мышечной сократимости.
Поверхность любой клетки, это фронт взаимодействия с внешней средой. Одни участки мембраны принимает энергию от внешней среды, другие, в преобразованном виде, отдают ее обратно. Активность единичной клетки, в масштабе многоклеточного организма, не эффективна, поэтому появляется необходимость в согласованном захвате и возвращении энергии группой клеток. На уровне нервно-мышечной системы, эта задача решается объединением специализированных нейронов в нервные узлы и мышечных элементов, в отдельные мышцы.

Кольцевые потоки импульсов, на суммарной мембране нейронной сети, уравновешивают поступивший из внешней среды избыток энергии, направляя его на активацию мышечной системы.
Одновременное поступление энергии в нервный узел, непосредственно контактирующий с внешней средой, изменяет мембрану его клеток, формируя барьер на пути внешнего воздействия, рецепторное поле. Группа клеток внутри нервной системы, не имеющая прямого контакта с внешней средой, принимает энергию от наружного нервного узла (для нее этот узел является своеобразной внешней средой). В конечном счете, цепочка нервных узлов образует канал поступления энергии от внешней среды. Одновременно развивается мышечная ткань, отдающая избыток энергии от нервного узла в окружающую среду.
Таким образом, в многоклеточном организме появляется ткань, поглощающая избыток энергии от окружающей среды и ткань, отдающая эту энергию обратно. Поскольку форма, в которой энергия поступает в нервную систему и выходит из нее, не совпадают, появляется избыток энергии, который расходуется усложнение нейронной сети и мышечной ткани. Одновременно усложняется система циклических потоков импульсов, оболочка, объединяющая нервную и мышечную ткани между собой.
Логично предположить, что нервная система развилась из клеточных элементов, которые непосредственно контактировали с мышечными клетками и с развитием нейронной сети, приобрела самостоятельное анатомическое представительство. Примером нейромышечной системы является нога улитки, имеющая свой рецепторный аппарат, реагирующий на изменения параметров внешней среды, проводящую систему и мышечную ткань, способную выводить избыток энергии из организма
7. Нервный узел, как система противодействия внешней среде
Поскольку избыток энергии, уравновешиваемый одной клеткой, невелик, в многоклеточном организме поглощение энергии осуществляется группой клеток, образующих функционально единый комплекс, нервный узел. В местах контакта нейронов с окружающей организм средой, где избыток энергии появляется особенно часто, происходит модификация клеточной оболочки с образованием рецепторов.

Наиболее простая модель нервного узла включает в себя тела нейронов, дендриты (их короткие отростки) и аксоны (длинные отростки нейронов следующего нервного узла). Внутри узла, упорядоченное объединение специализированных участков мембраны длинных и коротких отростков нейронов, формирует зону взаимодействий.
Через рецепторное поле, образованное аксонами нейронов первого нервного узла, в него поступает энергия из внешней среды. Поступившая энергия передается дендритами в зону взаимодействий, из которой она забирается аксонами следующего нервного узла (фактически, зона взаимодействий является внешней средой для аксонов следующего нервного узла). Нервный узел - это фильтр, структура которого частично уравновешивает внешнюю энергию. Избыток энергии, прошедший через первый нервный узел, уравновешивается структурами второго нервного узла и так далее. Таким образом, нервный узел можно рассматривать, как первый барьер на пути избытка энергии внешней среды.

Избыток энергии, неизбежно присутствующий в нервном узле, способствует формированию вставочных нейронов, образующих второй барьер на пути энергии, поступающей в нервный узел.
Поскольку объем прохождения энергии через нервный узел ограничен (в том числе и пропускной способностью следующего узла), в нем периодически возникает избыток энергии, который обуславливает дальнейшее развитие его структуры: увеличение количества нейронов, диаметра аксонов, числа и “ветвистости” дендритов. Со временем “структурный резерв” нервного узла исчерпывается и возникает необходимость в дополнительном барьере противодействия внешней среде. В нервном узле происходит появление вставочных нейронов, которые забирают на себя появившийся в узле переизбыток энергии и направляют его в зону взаимодействий, как тормозной медиатор.
Появление второго барьера противодействия внешней среде, соответствует феномену торможения в нервной системе. Если исходить из того, что нервный узел является фильтром, на пути избытка энергии, то легко можно представить логику появления тормозного медиатора. Зона взаимодействий является местом выхода энергии из нервного узла. Пресинаптическая мембрана дендритов нейронов выделяет в синоптическую щель возбуждающий медиатор. Если эффективность прохождения энергии через зону взаимодействий достигает максимума, возбуждающий медиатор становиться неэффективным и искажается процесс его выработки в окончаниях отдельных нейронов. Таким образом, возбуждающий медиатор вставочных нейронов преобразуется в свою функциональную противоположность, тормозной медиатор, который повышает порог чувствительности рецепторных полей зоны взаимодействий и частично нейтрализует избыток энергии. Тормозной медиатор, в нервных узлах расположенных внутри нейронной сети, использует избыток энергии на снижение эффективности возбуждающего медиатора, а в нервных узлах расположенных снаружи, на повышение порога чувствительности сенсорного анализатора.
Появление вставочных нейронов, второго барьера противодействия внешней среде, не исчерпывает все возможности нервного узла, как препятствия на пути избытка энергии. Возможно формирование третьего барьера противодействия внешней среде, что представляется следующим образом. Избыток энергии внутри нервного узла способствует разрастанию поверхности мембраны дендритов и появлению многочисленных ответвлений, образующих контакты с поверхностью нейронов данного узла, вне зоны взаимодействий. Образовавшаяся внутри узла сеть отростков получила название - нейропиль. К. Прибрам следующим образом характеризует особенности проведения импульсов в этой структуре (106):
“В нервных волокнах большого диаметра импульсы имеют, как правило, значительную амплитуду и быстро распространяются. В нервных волокнах с малым диаметром амплитуда электрического потенциала нервного импульса меньше, и скорость его проведения ниже. По мере удаления нервных волокон от тела клетки они утончаются и разветвляются: в результате этого импульсы становятся все более слабыми и замедленными, особенно в окончаниях волокон. Кроме того, более тонкие нервные волокна теряют толстое изолирующее покрытие (миелиновую оболочку), которая у крупных волокон препятствует взаимодействию импульсов. Медленнее потенциалы, возникающие в таких волокнах, не встречают препятствия для реального взаимодействия”
Таким образом, нейропиль представляет собой густую сеть отростков нейронов, помогающих рассеивать избыток энергии, проходящий по “основным магистралям” нервного узла. Прошедший нейропиль импульс, по своей количественной характеристике, соизмерим с энергией процессов, протекающих на мембране нейрона, поэтому он легко нейтрализуется энергией внутриклеточных структур. Мембрана нейрона, наподобие пружины, гасит рассеянный в нейропиле избыток энергии.
Как показал П.К. Анохин, попытка проследить смысл процессов, протекающих в нейропиле, с точки зрения проведения информации, неперспективна (7). В тоже время, появление нейропиля и его структура вполне оправданы, как механизм противодействия внешней среде. Импульсы, адаптированные нейропилем, достигая поверхности мембран клеток нервного узла, уравновешиваются силами внутриклеточных структур.

В нервном узле можно проследить схему трех барьеров противодействия внешней среде. Первый представлен самим нервным узлом, второй образован вставочными нейронами, вырабатывающими тормозной медиатор, третий связан с разрастанием нейропиля, рассеивающего избыток энергии по суммарной мембране нервного узла.
Несмотря на индивидуальные отличия отдельных нервных узлов, в каждом из них можно выявить три барьера противодействия внешней среде, что позволяет сделать следующие выводы:
Нервная система формируется как структура противодействия внешней среде, вход в которую осуществляется через рецепторы, а выход, через структуры, вырабатывающие медиатор
Нервная система состоит из цепочек нервных узлов, объединенных в совместную нейронную сеть, которые являются каналами проведения энергии поступившей через рецепторное поле
В процессе развития нервного узла в нем формируются первый барьер противодействия внешней среде - увеличение количества нейронов, рост аксонов, дендритов и образование между ними зоны взаимодействий
Торможение, второй барьер на пути внешних воздействий, обусловлено формированием в нервном узле вставочных нейронов, ограничивающих эффективность прохождения энергии через зону взаимодействий
Третий барьер противодействия внешней среде обеспечивает нейропиль, который рассеивает избыток энергии по суммарной мембране нервного узла, вне зоны взаимодействий
8. Схема эволюции нейронной сети
В монографии К. Прибрама, в разделе, где он рассматривает проблемы запоминания, высказана идея о том, что отдельные участки нервной системы напоминают голографическую пластину. В основном речь идет о системе нейропиля, структуре характерной для большинства нервных узлов. По его мнению, процессы запоминания в нервной системе формируются на мембране нейронов, при контакте с ними медленных потенциалов, возникающих в нейропиле и имеющих волновой характер (106). Верная, с нашей точки зрения, гипотеза о том, что запоминание в нервной системе процесс, родственный таковому при записи и воспроизведении голографических образов, используется несколько односторонне. Согласно нашим представлениям, процесс развития нервной системы, на всех его этапах, напоминает формирование голографической пластины. Каждое образование нейронной сети, каждый ее этаж, уравновешивает набор циклически организованных фрагментов внешнего мира. Изменения внешней среды нарушают равновесие с нервной системой, что порождает динамику ее состояний, а содержание сознания, в связи с этим, приобретает подвижность (нечто подобное происходит при последовательном проецировании, с определенной скоростью, неподвижных кадров фильма на экран). Попробуем представить нейронную сеть и окружающую ее среду, в качестве двух динамических систем, которые уравновешивают друг друга.

Три уровня организации материи получают свое отражение в трех, последовательно образующихся, этажах нейронной сети.
На схеме, в окружающей нервную систему среде, показаны устойчивые элементы, которые образуют несколько уровней движения. Взаимодействие циклических систем первого уровня формирует второй уровень движения, который в свою очередь, входит в состав циклической формы движения третьего уровня. Нервная система представлена на схеме в виде трех, последовательно образованных этажей, где участки “А” принимают из окружающей ее среды избыток энергии, а участки “Э”, отдают его обратно.
Появление первого слоя нервных узлов уравновешивает избыток энергии, возникающий в первом уровне организации материи (энергия этого уровня способна преодолеть порог чувствительности сенсорного анализатора и придать потокам импульсов, в наружном слое нервных узлов, логику перемещения элементов, первого уровня внешней среды). Второй слой нервной системы, не имея прямого выхода в окружающую среду, реагирует на неуравновешенные формы движения в первом слое, поэтому дальнейшее взаимодействие, между нервной системой и внешней средой, уравновешивает избыток энергии, отражающий активность второго и третьего уровня организации материи, что выражается в появлении соответствующих слоев нервной системы.
Логика рассуждений показывает, что одновременная активность всех слоев нервной системы формирует относительно точную динамическую копию окружающего мира, при этом первый слой нервной системы отражает активность определенного уровня движения внешней среды, а последующие слои уравновешивают уровни движения, которые сформировались на базе элементов этого уровня. Чем дальше расположен очередной слой нервной системы от первого слоя, тем более абстрактный образ окружающего мира в нем формируется.
В первую очередь, данные рассуждения справедливы для сенсорной части нервной системы, куда поступает энергия внешней среды. В тоже время, выход энергии, который осуществляется моторной частью нервной системы, имеет определенные закономерности. Вначале активность мышечной системы хаотична и ее единственным результатом является вывод энергии из нервной системы. Со временем, те сочетания мышечной активности, которые кратно повторяются и приводят к равновесию с внешней средой, запоминаются (моторная часть нейронной сети последовательно уравновешивает активность отдельных слоев мышечной системы, в результате чего происходит “многослойная запись” поведения организма). Таким образом, формирование моторной части нейронной сети происходит по аналогии с формированием ее сенсорной части (различие между частями нейронной сети условное, и определяется особенностями динамических систем, с которыми они образуют равновесие). Следовательно, в нервной системе, по мере ее развития, образуется динамическая копия сред, с которыми она контактирует, в том числе и модель поведения организма, которая уравновешивает определенные процессы в окружающей среде. В конечном счете, приходим к выводу, что в нервной системе образуется поле сознания, оболочка, которая по мере своего развития, уравновешивает растущее количество факторов внешней среды. Поступление внешней энергии в эту оболочку происходит через сенсорную часть нейронной сети, а выход, через моторную часть.
9. Схема эволюции моторной части нейронной сети
В наиболее общем виде, структуру моторной нейронной сети можно представить следующим образом. Наружный слой моторной сети непосредственно контактирует с отдельными мышцами. Поскольку мышечная система выводит избыток энергии из мозга, в нервных узлах, напрямую с ней связанных, отсутствует избыток энергии и не образуется система вставочных нейронов (второй барьер противодействия внешней среде). Между узлами первого слоя моторной сети и отдельными элементами мышечной ткани образуется кольцевой поток импульсов, количественная сторона которого отражает уровень активации того или иного участка мышечной системы. Подобное рефлекторное кольцо способно принимать избыток энергии от контактирующих с ним участков нервной системы и преобразовывать его в мышечную активность. Поступающая в моторный узел энергия, последовательно повышает уровень пульсации афферентных и эфферентных нейронов, после чего расходуется на активацию мышечных элементов (рефлекторное кольцо можно сравнить с насосом, отдающим в окружающую среду избыток энергии, поступающий из рядом расположенных отделов нервной системы). Количество нейронов данного узла и пропускная способность их отростков, определяется массой мышцы, с которой они контактируют.
Избыток энергии в сенсорных узлах, который не может поглотиться моторными узлами первого слоя, в виду их ограниченной пропускной способности, способствует увеличению количества моторных нейронов, что приводит к формированию второго слоя моторных узлов. Узел второго слоя моторных узлов, связан с несколькими узлами первого слоя, отражающих активность функционально идентичных мышечных элементов, которые могут находиться в антагонистичных отношениях друг к другу. В виду невозможности одновременного сокращения мышц, в моторных узлах второго слоя появляется избыток энергии, который может быть использован на формирование вставочных нейронов (второго барьера противодействия внешней среде).
Формирование вставочных нейронов в моторных узлах второго слоя происходит следующим образом. В момент, когда одна из мышц уже поглотила энергию и сократилась, связанные с ней афферентные нейроны не могут передать в зону взаимодействий поступающий в них избыток энергии. Этот избыток способствует переориентации части афферентных нейронов, на вставочные, которые передают поступивший в них избыток энергии не только третьему слою нервных узлов, но и той части нейронов данного узла, которые связаны с расслабленной мышцей-антагонистом. Поскольку мышца-антагонист не может сокращаться одновременно со своим антиподом, происходит искажение механизма выработки медиатора вставочных нейронов, которые начинают выделять вместо возбуждающего медиатора тормозной, направляя его в зону взаимодействий, связанную с мышцей-антагонистом (следует подчеркнуть, что логика развития нервного узла определяется логикой оптимального распределения в нем избытка энергии).

Динамическая система импульсов, в нервных узлах первого слоя, уравновешивает энергию, возникающую при сокращении связанных с ними мышц. Во втором слое уравновешивается энергия, возникающая при попеременной активности нервных узлов предыдущего слоя. Таким образом, избыток энергии, поступающий во второй слой моторных узлов, при участии вставочных нейронов, активирует упорядоченную мышечную активность (программы поведения).
Циркулярные потоки импульсов, в первом слое моторных узлов, уравновешивают сокращения и расслабления мышечных единиц, находящихся в одной стадии активности. В моторных узлах второго слоя появляются две группы нейронов, которые находятся в противоположном энергетическом состоянии, что закрепляется тормозными элементами. Такие нервные узлы уравновешивают попеременную активность мышц антагонистов. В третьем и последующих слоях моторной нейронной сети, потоки импульсов уравновешивают объединения, в состав которых входит несколько мышц, находящихся в антагонистических отношениях. Таким образом, поступление избытка энергии на верхние этажи моторной сети способно активировать движение мышц любой сложности. Ттношения.ульсов уравновешивают объединения из нескольких мышц, находящихся в антогонистических

Развитие моторной рефлекторной сети уравновешивает все формы активности мышечной системы, поэтому процесс становления равновесия является, по своей сути, запоминанием соответствующих программ поведения.
Важно подчеркнуть, что развитие нервной системы определяется законами становления равновесия между двумя динамическими системами и не нуждается во внешнем управлении.
10. Схема формирования сенсорной нейронной сети
Развитие сенсорной нейронной сети, в наиболее общем виде можно представить, как последовательное развитие многослойной структуры, каждый слой которой уравновешивает определенные факторы внешней среды. Схематически взаимоотношения нейронной сети и внешней среды можно изобразить в виде двух динамических систем, стремящихся к равновесию:

Избыток энергии, периодически возникающий внешней среде, формирует в нервной системе уравновешивающую его цепочку нервных узлов.
Избыток энергии, периодически возникающий в окружающей обстановке, вызывает появление и развитие сенсорной нейронной сети. В местах контакта нейронов с внешней средой появляются рецепторы, формирующие рецепторное поле данного нервного узла. Параллельно происходит увеличение количества нейронов и развитие их отростков. Воздействие внешней среды на рецепторное поле способствует появлению в нервном узле определенного сочетания активности нейронов, характеризуемое как узор информации. Избыточная энергия, не успевающая выйти из нервного узла, определяет появление вставочных нейронов, вырабатывающих тормозной медиатор, который ограничивает эффективность прохождения импульсов в зоне взаимодействий (на входе в нервный узел).
Поступивший через рецепторное поле узор информации, продвигается по цепочке взаимосвязанных нервных узлов, способствуя появлению в них родственных узоров информации. Каждый нервный узел, является барьером на пути внешнего воздействия и отфильтровывает часть внешней энергии, поэтому динамическая модель внешней среды, в удаленных слоях нейронной сети, содержит упрощенный (абстрагированный) образ реальности. Процесс уравновешивания энергии внешней среды, изменяет структуру нейронной сети и является процессом консолидации следов памяти в нервной системе, что будет обсуждаться выше.
В своих работах, посвященных физиологии зрительного анализатора, Д. Хьюбел показал, что поступивший через рецепторное поле импульс энергии, не только проходит через цепочку взаимосвязанных нервных узлов, но и расходуется на появление зоны торможения вокруг фокусов входа энергии (131). Как видим, данная информация полностью совпадает с нашей теоретической моделью сенсорного анализатора и представлением о нервных узлах, как барьерах противодействия внешней среде. Узор информации, прошедший через рецепторное поле, несет в себе логику движения внешней среды, которая проявляется в характере циркуляции импульсов во всех слоях сенсорного анализатора (процесс абстрагирования свойственен любой системе последовательно собранных фильтров, поэтому о сенсорном анализаторе можно говорить, как об автоматическом “обобщающем устройстве”, вырабатывающем “абстрактные” образы внешнего мира).

Энергия внешнего воздействия уравновешивается перестройкой характера кольцевого движения импульсов в нервном узле, что в свою очередь вызывает изменение его структуры и структуры, связанных с ним нервных узлов.
Важно обратить внимание, что появление в системе зрительного анализатора родственных узоров информации, возможно только вследствие изменения параметров внешней среды. Показателен в этом отношении опыт по обездвиживанию изображения на сетчатке глаза, посредством проецирования изображения на экран через зеркальце, укрепленное на склере исследуемого субъекта. При обездвиживании изображения на сетчатке глаза происходит исчезновение изображения на экране (93). Это лишний раз подтверждает тот факт, что образы сознания объективны, так как являются продуктом движения внешней среды (ранее обсуждалось, что объективная информация о внешнем мире заключена в движении).
В свое время, Г. Гельмгольц, изучая физиологию зрения, пришел к логичному выводу, что ощущения представляют собой лишь знаки, “иероглифы”, не имеющие никакого сходства с реальными свойствами вещей (126). Спорить с этим трудно. Форма и качество объектов окружающего нас мира относительны. В тоже время, факт их перемещения объективен и содержит независимую от нас информацию. То или иное событие, можно описать в книге, изобразить на картине, снять в кино и так далее. Несмотря на то, что в каждом варианте изложения применялись непохожие друг на друга “иероглифы”, логика их перемещения узнаваема и позволяет утверждать, что речь идет об одном и том же событии.

Появление в цепочке узлов сенсорного анализатора родственных, то есть совместимых друг с другом узоров информации, способствует формированию циклических потоков импульсов между соседними узлами внутри самого анализатора.
В структуре нервного узла предусмотрено три основных механизма противодействия избытку энергии, поступающему из внешней среды. Однако процесс нейтрализации избытка энергии требует дополнительных условий, которые формируются по мере развития нейронной сети. Механизмами нейтрализации избытка энергии в нейронной сети являются: развитие ее инфраструктуры, формирование совместных зон, через которые энергия может поступать из сенсорных отделов в моторные отделы нейронной сети и формирование “вставочных” нервных узлов, обеспечивающих циркуляцию “дежурного” узора информации внутри сенсорного анализатора.
Д. Хебб обратил внимание на существование в нервной системе колец реверберации, наличием которых он объяснял механизмы кратковременной памяти (93). Кольцевые потоки импульсов в нервной системе, Г. Шеперд и К. Прибрам рассматривали в контексте обратной связи (141, 106). По нашему мнению, в нейронной сети, под воздействием избытка энергии, образуется аналог вставочных нейронов – вставочные узлы. Благодаря родственному характеру узоров информации, поступающему в цепочку взаимосвязанных нервных узлов, возможно формирование кольцевых потоков импульсов, между элементами цепочки, где часть энергии внешней среды возвращается в предыдущий слой нейронной сети. Поскольку в нервных узлах, внутри нейронной сети, не имеет смысла ограничивать поступление энергии на входе, вставочные узлы вырабатывают возбуждающий медиатор. В процессе кольцевого движения потока импульсов происходит рассеивание избыточной энергии. Такая кольцевая структура, образованная потоком импульсов, устойчивая в пространстве и во времени, является материальной основой образов в нашем сознании. Как видим, феномен обратной связи не есть проявление каких-то таинственных сил, контролирующих поступление информации в нервную систему. Он является субъективным восприятием циклической формы движения, лежащей в основе организации материи. Рассеивание избытка энергии, в процессе прохождения по кольцу, это барьер противодействия внешней среде, образовавшийся на уровне нейронной сети.
Согласно данным, которые опубликовал Д. Хьюбел, кора зрительного анализатора имеет многослойное строение. Нейроны каждого слоя специализируются на восприятии определенной зрительной информации (Д. Хьюбел предложил называть их нейронами-детекторами, соответствующих признаков). Данный факт ввел в недоумение многих исследователей нервной системы, так как не укладывался в существующие теоретические построения (131). С точки зрения излагаемого материала, появление нейронов-детекторов вполне логично. Каждый слой стриарной коры представляет собой развернутый на плоскости нервный узел. Для аксонов нейронов верхнего слоя коры предыдущий слой является внешней средой, при контакте с которой они реагируют на тот или иной вид избытка энергии (тоже происходит при формировании специфического рецепторного аппарата сенсорного анализатора). Система вставочных нейронов, в пределах одного слоя коры, обеспечивает стабильное существование узора информации, характерного для данного уровня равновесия кольцевых потоков импульсов с внешней средой.
Поступающие из внешней среды волны энергии образуют в нервной системе циклические структуры, устойчивые в пространстве и времени. С точки зрения циклической организации материи, они являются “материальным телом” определенного уровня движения. Подобные “тела” субъективно воспринимаются образами “мира внутри нас”. Чем сложнее устроена нервная система, тем больше в ней слоев и тем устойчивей, в плане зависимости от внешней среды, будут в ней образы нашего сознания. Таким образом, в нейронной сети можно выделить три барьера противодействия избытку энергии, поступающему из внешней среды:
Первый барьер обусловлен ростом самой сети и формированием дополнительных уровней ее организации
Второй барьер обеспечивается выходом избытка энергии, через совместные участки, в моторную часть нейронной сети, а оттуда во внешнюю среду, в виде мышечной активности
Третий барьер обеспечивается появлением вставочных узлов, преобразующих последовательное продвижение импульсов, по цепочке родственных нервных узлов, в систему взаимосвязанных друг с другом кольцевых потоков импульсов
11. Принципы взаимодействия сенсорного и моторного отделов в нейронной сети
Существование сенсорной части нейронной сети не имеет смысла, если она не будет отдавать поступающую в нее энергию во внешнюю среду. Факт того, что некоторая часть внешней энергии уравновешивается инфраструктурой мозга, не отличает нервную ткань от прочих тканей организма. Только в единстве сенсорной моторной частей нейронной сети, наряду с наличием мышечной ткани делает возможным существование уровня движения, образованного кольцевыми потоками импульсов, поля сознания. В процессе изучения нервной системы морского моллюска Aplysia, Э. Кэндел установил наличие в ней командных нейронов, активность которых запускала определенные программы поведения, с участием множества соподчиненных нейронов (70). Такие нейроны имели связь с сенсорными образованиями в нервной системе, что позволяло им активировать программу поведения в ответ на определенную внешнюю ситуацию. В мозге человека существуют обширные области коры, в которых объединяются моторные и сенсорные участки нервной системы. Совместные образования определяются во всех анатомических отделах мозга (стволе, подкорковой области, коре).
Таким образом, в нейронной сети существуют участки, общие для сенсорной и моторной частей нервной системы, через которые избыточная энергия поступает в моторный мозг, где существуют условия для ее выхода в окружающую среду. Логика образования каналов вывода энергии представляется следующим образом. В сенсорном мозге, по мере его эволюции, появляется система вставочных узлов, которые разбивают линейный канал движения импульсов в нейронной сети, на взаимосвязанные, кольцевые структуры. В тех участках нейронной сети, где поток импульсов застаивается, появляется избыток энергии и происходит увеличение массы нейронов. В конечном счете, вставочный узел разбивается на части и происходит образование зоны взаимодействий, из которой формируется каналы вывода энергии. Один из участков вставочного узла, образует канал выхода в зону взаимодействий узла, находящегося в моторной части нервной системы, что активирует сокращение мышцы и способствует выходу энергии. По аналогии с командными нейронами, описанными Э. Кэнделом, видоизмененные вставочные узлы можно назвать командными. Командные узлы вырабатывают возбуждающий медиатор, поступающий в моторные узлы (тормозной медиатор не может быть эффективным в контексте создавшейся ситуации). Следует подчеркнуть, что избыток энергии не всегда находит выход в моторной сети, особо, если он поступает в ее верхние этажи, не имеющие непосредственного контакта с мышечной тканью. Создавшийся избыток энергии способствует усложнению моторной сети, а вместе с ней и поведения организма.

Цепочка сенсорных узлов берет энергию из внешней среды. Командный узел образует кольцо обратной связи между сенсорными узлами второго - первого слоя и афферентной зоной моторного узла. Из зоны взаимодействий командного узла, возможно отведение избытка энергии в зону взаимодействий моторного узла.
Поток импульсов может быть стабильным в нейронной сети только при их кольцевом движении (механизм и целесообразность обратной связи обсуждались выше). Круговое продвижение потока импульсов, имеющего определенную структуру (узор информации), способствует развитию в нейропиля, который в свою очередь модифицирует структуру суммарной мембраны нервных узлов. Особо выражены эти процессы в коре мозга, каждый слой которой является нервным узлом.
Как видим, благодаря кольцевому продвижению импульсов, в командном узле постоянно присутствует узор информации, который состоит из совмещенных узоров от сенсорного и моторного узлов. Раздражение такого узла одновременно порождает сенсорный образ и программу поведения. Схему объединения сенсорной и моторной частей нейронной сети, через командные узлы, можно изобразить в упрощенном виде. Это позволит яснее представить, из каких модулей состоит нервная система.
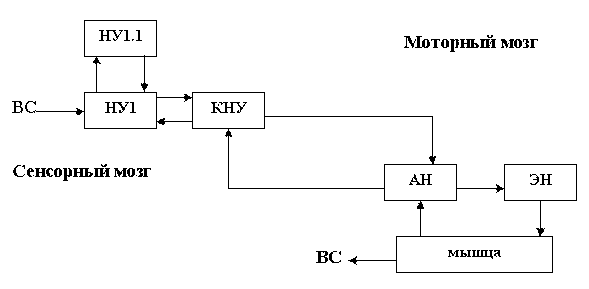
Принципиальное устройство совместного участка моторной и сенсорной нейронных сетей.
Принципы строения нейронной сети применимы к организации коры мозга. Поток энергии поступает не только в средний слой коры, который обеспечивает его продвижение в соседние участки, но и в нижерасположенные слои, которые образуют кольцо обратной связи. Отдельным нервным узлам, так же как и отдельным нейронам, из которых они состоят, свойственна пластичность. В случае неэффективности возбуждающего медиатора, окончания их отростков, в процессе эволюции нейронной сети, может перестроиться к выработке тормозного медиатор, что повышает возможности утилизации избытка энергии (химическая структура тормозного и возбуждающего медиаторы во многом идентичны).
Сенсорный и моторный мозг составляют единую систему, стремящуюся к равновесию с окружающей средой. В сенсорном мозге отражаются параметры внешней среды, а в моторном отражаются программы воздействия на нее. Становление равновесия между сенсорной и моторной частью нервной системы, является фрагментом обмена энергией между организмом и средой (мышца, совершая движение “отсасывает” энергию из нервной системы, а в освободившуюся нишу поступает избыток энергии внешней среды, который, будучи пропущен через фильтр сенсорного мозга, видоизменен).
Оболочка, состоящая из циклических потоков импульсов, оказывает постоянное давление на суммарную мембрану нейронов мозга, заставляя их видоизменяться. Потенциальная энергия, консолидированная в измененной структуре нервной системы, составляет основу памяти. Чем сложнее эта структура, тем организованней становиться процесс становления равновесия с внешней средой. Например, выход энергии из нервной системы обеспечивается хаотическими движениями. В тех случаях, когда движение случайно уравновешивает воздействие внешней среды, поступление энергии в нервную систему прекращается и кольцевые потоки импульсов, содержащие определенный узор информации, стабилизируются. Продвижение измененного узора информации по кольцу уравновешивается мембраной нейронов, деформируя ее (процесс консолидации). Кратность возникновений состояний равновесия, с теми или иными параметрами среды, определяет особенности развития всех макро и микроструктур мозга. Поскольку изменения происходят параллельно, в сенсорном и моторном мозге возникает феномен ассоциирования, объединение узоров сенсорной и моторной информации в едином узоре поля сознания.
Хаотическая форма движения является основной формой поведения, для примитивно организованных организмов и выполняет функции насоса, откачивающего энергию из их нервной системы. Со временем, случайные результаты усложняют структуру моторного мозга и соответственно рисунок хаотических движений. Появляются универсальные приемы поведения в нестандартных ситуациях, которые составляют основу ориентировочного рефлекса, который, в свою очередь, лежит в основании формирования всех прочих форм поведения. Наивысшей формой развития ориентировочного рефлекса является мышление, процесс который стихийно вырабатывает и закрепляет новые программы поведения. Определенные, стихийно закрепившиеся закономерности структуры ориентировочного рефлекса определяет вектор движения нервных процессов, и отражают взаимодействие организма с внешней средой, что составляет суть феномена “логика мышления”.
В свое время Н.А.Бернштейн предположил, что в моторной коре мозга представлена топологическая проекция пространства. Отрицательное отношение к данной идее высказал К. Прибрам (106). Согласно его мнению, в моторном мозге присутствуют только “силы возбуждающие мышечные рецепторы”. Насколько был прав Н.А. Бернштейн, становиться ясным по ходу нашего изложения. Формирование поведенческого континуума происходит “на ощупь”, методом проб и ошибок. Установившееся равновесие с внешним миром подразумевает наличие в моторном мозге его негативной копии. Н.А. Бернштейн интуитивно чувствовал, что сенсорная и моторная части нервной системы составляют единое целое и их надо рассматривать совместно (20):
“… в организме все моторы осенсорены, все сенсоры омоторены”
В нашем понимании, оболочка из циклических потоков импульсов образует уровень движения на совместной мембране нейронов всей нервной системы, который уравновешивает определенные проявления внешнего мира и становиться его динамической копией. В рамках единой динамической системы, разделение потока импульсов, на сенсорную и моторную части, относительно. Подобно жидкости, циклически организованные импульсы, формирующие поле сознания, принимают форму того сосуда, в котором они находятся.
12. Память как следствие движения
К. Лешли, в одной из работ, посвященной проблемам памяти, высказал следующее мнение (106):
“Анализируя данные, касающиеся локализации следов памяти, я испытываю иногда необходимость сделать вывод, что обучение вообще невозможно. Тем не менее, несмотря на такой довод против него, обучение иногда происходит”
Проблему памяти обычно рассматривают, как частный случай той или иной области знания, избегая более широких обобщений. Например, в монографии И.П. Ашмарина, в которой рассматривается проблема хранения информации на биологическом уровне, предложено следующее определение (16):
“Биологическую память можно определить, как способность живых
существ (или их популяций), воспринимая воздействия из вне, закреплять, сохранять и в последующем воспроизводить вызываемые этими воздействиями изменения функционального состояния и структуры.”
Далее, развивая свою точку зрения, И.П. Ашмарин обращает внимание на такие разновидности памяти, как эпигенетическая, иммунологическая и нейробиологическая. Если сравнить между собой различные определения памяти, то можно выявить общие закономерности:
Поскольку память, это динамический процесс, то обсуждать проблему памяти имеет смысл в том случае, если что-то запоминается и что-то воспроизводиться.
В процессе запоминания мы выделяем две фазы, хранения и воспроизведения, которые периодически сменяют друг друга, что позволяет говорить о циклической форме движения
В контексте философии движения, память представляется, как субъективное представление об универсальном свойстве материи, процессе колебания энергии, между исследуемой системой и окружающей средой. Например, вода, отдавая энергию, при температуре ниже нуля “вспоминает”, что ей надо замерзнуть, а при температуре выше нуля, поглощая энергию, “воспроизводит” свое исходное состояние. Можно привести бесконечное множество примеров “воспроизведения” в “неживой природе”, когда объект, помещенный в определенные условия, регулярно проявляет те или иные свои свойства. Традиционное использование понятия “память”, в пределах той или иной области знаний, связано с употреблением узко специальной терминологии, что вуалирует сущность процесса и препятствует более широким обобщениям. Мы можем предложить следующее определение памяти, как одного из атрибутных свойств материи:
Память является процессом становления равновесия, между условно выбранной системой и окружающей ее средой. Особенности строения системы, определяют феномен “запоминания”, а феномен “воспроизведения”, это субъективное представление о форме, в которой происходит выравнивание нарушенного равновесия.
Когда мы говорим о запоминании, то имеем в виду хранение энергии в потенциальной форме, которая, в определенных условиях, способна произвести то или иное действие. Такой подход подразумевает существование двухфазного процесса. Фаза поглощения энергии, соответствует ее переходу в потенциальное состояние, а фаза освобождения совпадает с превращением потенциальной энергии, в кинетическую энергию. Субъективно, обе фазы могут восприниматься раздельно, как не связанные друг с другом процессы или как единое целое, следствие поглощения энергии системой. Например, фазы перемещения маятника часов, трудно отнести к феномену памяти в привычном для нас понимании. Однако, происходящей при этом процесс превращения кинетической энергии в потенциальную энергию, мало отличается по своей сути от процессов, которые мы характеризуем, как запоминание и воспроизведение. Если бы мы не видели маятника часов, а наблюдали лишь следствия его движения, то могли бы оценивать происходящее, как феномен запоминания и воспроизведения чего-либо в той или иной среде. Таким образом, память может быть представлена, как процесс превращения энергии в веществе:
Память, это субъективное представление о колебании энергии в системе, где фаза воспроизведении соответствует освобождению кинетической энергии, а фаза запоминания, ее переходу в потенциальное состояние
С точки зрения биологии, память, это потенциальная энергия, содержащаяся в системе, которая заложена на различных уровнях организации организма, генетическом, клеточном, тканевом и так далее. Выделение того или иного вида памяти, зависит от условий эксперимента и логической интерпретации полученных результатов. Так представление о генетической памяти, обусловлено исследованием процессов протекающих в клетках на молекулярном уровне, изучение биохимических особенностей внутренней среды организма, порождает определение иммунологической памяти и так далее.
Нейробиологическую память определяют колебательные взаимоотношения вещества мозга с окружающей средой. Постоянные внешние воздействия способствуют появлению в нервной системе своеобразных “пружин”, барьеров противодействия окружающей среде, которые, при повторном соприкосновении организма с данными условиями среды, распрямляются, восстанавливая утраченное равновесие, что субъективно воспринимается, как фаза воспроизведения. Очевидность подобного подхода, к рассмотрению памяти, вуалируется тем, что мозг в целом не воспринимается, как динамическая система и процесс его противодействия изменениям внешней среды, рассматривается в изолированном виде, без понимания общей направленности всех процессов в нем происходящих.
При рассмотрении мозга, как динамической структуры, в нем можно выделить нейронную сеть, в которой существует множество циклически организованных потоков импульсов, образующих поле сознания, систему устойчивую в пространстве и времени (поле сознания можно сравнить с водой в сосуде, любое воздействие на которую, вызывает волну изменений, распределяющуюся по всей системе). В нервной системе, циклические потоки импульсов можно рассматривать, как своеобразный буфер, между двумя стремящимися к равновесию системами, окружающей средой и телами нейронов. Поскольку постоянного равновесия между этими системами не может быть, в поле сознания периодически появляется избыток энергии, который изменяет структуру нейронной сети, временно восстанавливая утраченное равновесие.
Периодические изменения макро и микроструктуры нейронной сети (оправы в которую заключено поле сознания), являются основой долговременной памяти. Кратковременная память соответствует состоянию временно нарушенного равновесия в поле сознания. Если перестройка сознания, при изменении окружающей среды, происходит при участии ранее сформировавшихся изменений мембраны нейронов, мы говорим о долговременной памяти. Если мембрана не имеет механизма противодействия изменившимся условиям внешней среды, происходит ее перестройка, то есть обновление долговременной памяти. Таким образом:
Кратковременная память, это состояние временно нарушенного равновесия между полем сознания и внешней средой, которое не уравновешено ранее появившимися изменениями мембраны нейронной сети и требует ее перестройки
Анатомическая эволюция нервной системы (увеличение числа нейронов, развитие проводящих путей и прочее), происходящие при эволюции нервной системы, составляют основу филогенетической памяти . Прижизненные изменения мембраны нейронов (развитие нейропиля, пластические изменения пресинаптических и постсинаптических мембран), обуславливают способность нервной системы запоминать и воспроизводить текущие внешние воздействия, составляют основу памяти мембраны (именно она трактуется, как память в общепринятом значении). Промежуточное положение, между этими видами, занимает память онтогенетическая (в онтогенезе происходит созревание нервной системы, проявляющееся увеличением количества нейронов, ростом окончаний их отростков, а в дальнейшем, изменениями микроструктуры мембраны нейронов, в местах синаптических контактов). Таким образом:
Сознание – это своеобразная оболочка, динамическая структура, заключенная в оправу нервной ткани, уравновешивающая определенные состояния внешней среды и тканей организма
Память нервной системы заключена в изменениях суммарной мембраны нейронов, которые, согласно ранее приобретенного опыта, модулируют ее взаимоотношения с внешней средой
Фаза запоминания, это перестройка ряда участков на суммарной мембране нейронов нервной системы, происходящих вследствие изменений внешней среды, нарушающих равновесие процессов протекающих в сознании
Фаза воспроизведения, это противодействие внешним воздействиям со стороны ранее изменившихся участков мембраны нейронов, что определяет соответствующее содержание сознания
13. Феномен заполнения в движущейся системе
Любая система, находящаяся в состоянии динамического равновесия с окружающей ее средой, обладает определенной устойчивостью. Устойчивость системы определяется объемом энергии внешней среды, который она способна поглощать и возвращать обратно, в преобразованном виде, сохраняя логику своей организации. Поступившая в систему энергия расходуется на последовательное увеличение скорости циклического движения отдельных ее элементов. Продвигаясь от цикла к циклу, энергия выходит в окружающую среду, при этом структура самого материального тела сохраняется. Например, многие объекты, изменяя свои свойства при нагревании, способны их восстановить в процессе остывания, при этом избыток тепловой энергии, в виде волнового процесса, рассеивается в соседних системах. Если поступившая в систему энергия вызывает в ней необратимые изменения, происходит ее положительная или отрицательная эволюция, с последующим восстановлением утраченного равновесия.
Процессы запоминания в нервной системе происходят при поступлении избыточной энергии, в количестве, достаточном для изменения ее структуры (в результате чего устанавливается равновесие с внешней средой на более высоком уровне). Если избыток энергии, поступивший в нервную систему, уравновешивается ее структурами и не вызывает их эволюцию, мы говорим, применительно к памяти, о процессах воспроизведения (уравновешивание избыточной энергии происходит вследствие ранее приобретенной устойчивости мембраны нейронной сети к данному воздействию). В этом контексте можно предположить существование двух видов воспроизведения. В первом варианте, одни и те же участки мембраны нейронов реагируют на некое множество близких по содержанию состояний внешней среды. Во втором варианте, когда внешние условия содержат элемент новизны, равновесие обеспечивается необычным сочетанием активности различных участков мембраны, без нарушения ее структуры (то есть, впервые возникшее состояние среды уравновешивается уже существующими средствами).
Поскольку на целый ряд внешних условий нервная система реагирует без перестройки мембраны нейронов одними и теми же механизмами, или необычным их сочетанием, (то есть без расширения резервов памяти), процесс воспоминания можно рассматривать, как “феномен заполнения”.
Феномен заполнения, это процесс выравнивания равновесия с внешней средой, при котором рассматриваемая система качественно не изменяется
Таким образом, подчеркивается, что воспоминание не всегда есть воспроизведение событий прошлого и в ряде эпизодов мы будем иметь совершенно новые состояния сознания, ничего общего не имеющие с процессом воспроизведения информации.
Чем сложнее система, тем совершенней ее способность уравновешивать похожие друг на друга состояния внешней среды одними и теми же механизмами или их необычными сочетаниями. Относящиеся к запоминанию или научению процессы, это частный случай феномена заполнения, проявления которого охватывают значительно более широкий круг явлений. Например, И.П.Павлов, занимаясь изучением условных и безусловных рефлексов, в различных условиях окружающей среды, по сути дела имел дело с феноменом заполнения. Повторение условной и безусловной ситуации образует динамическое равновесие внешней среды и нервной системы. Если воспроизведение условной ситуации происходит с небольшим дефектом, отсутствие безусловного раздражителя, имеет место феномен заполнения и система реагирует стандартными, ранее сформированными механизмами.
Представление о феномене заполнения объясняет ряд парадоксов, которые не укладывались в концепцию воспроизведения содержимого памяти. Проявления феномена заполнения имеют место при спонтанном воспоминании, логично не связанном с конкретной обстановкой, в которой оно появилось. К данному классу явлений следует относить интуитивное решение той или иной проблемы, “фантом ампутированных”, у людей потерявших одну конечность (45), способность живых организмов к регенерации (самовосстановление поврежденной клетки, восстановление дефекта тканей организма, после ранения и так далее). Когнитивные процессы (феномен ассоциирования), тоже следует рассматривать в контексте феномена заполнения.
В плане излагаемого материала следует обратить внимание на то, что процессы запоминания и воспроизведения имеют определенную направленность, стремление обрести равновесие с окружающей средой.
14. Филогенетическая память нервной системы
Когда специалисты по психологии или нейрофизиологии характеризуют тот или иной вид памяти, они часто опускают из виду, что она представляет собой нечто целое, свойственное всей нервной системе. Фактически, память это состояние сознания, оболочки состоящей их кольцевых потоков импульсов на суммарной мембране нейронов всей нервной системы, которая реагирует на любые внешние воздействия целиком, а не отдельными изолированными частями. Структурная организация этой оболочки определяется равновесием между параметрами внешней среды и структурой самой нейронной сети (все анатомические и функциональные образования в мозге имеют непосредственное отношение к ее формированию).
Суммарная мембрана нейронной сети создает оправу, придающую форму потоку импульсов. В тоже время, под давлением этого потока происходит ее деформация, причем процесс этот неравномерный и зависит от особенностей взаимоотношения входных рецепторов с окружающей средой (деформацию мембраны можно сравнить с изменениями рельефа местности, под воздействием сил природы). Характеризуя память, как процесс, следует обратить внимание на ее медленный компонент, обусловленный анатомическими особенностями организации мозга, сложившимися в процессе эволюции и быстрый компонент, обусловленный прижизненной пластичностью суммарной мембраны нервной системы. Медленный компонент памяти характеризуется увеличением количества нейронов, разрастанием поверхности их мембраны, появлением проводящих путей и нервных узлов, а быстрый – изменением локальной структуры мембраны, в местах синаптических контактов.
Динамика процесса эволюции нервной системы недоступна для нашего непосредственного восприятия и оценивается лишь ретроспективно, в материалах по ее сравнительной эволюции. Формирование макроструктур в нервной системе можно рассматривать, как фазу “запоминания” (консолидации) воздействий среды на протяжении развития данного биологического вида. Отдельные образования в мозге отражают те или иные уровни равновесия между внешней средой или тканями организма (фактически, волновой процесс на суммарной мембране нейронов, это фронт взаимодействия тканей организма с окружающей средой, переведенный на язык движения нервных импульсов). Усложнение нервной системы выражается в появлении барьеров противодействия, которые уравновешивает все большее количество вариантов изменений внешней среды. Следовательно:
Структурная организация нервной системы может рассматриваться в качестве филогенетической памяти, скорость формирования которой соизмеримо со скоростью развития биологического вида
С точки зрения излагаемого материала, представление о том, что анатомические образования в нервной системе это филогенетическая память, вполне логично. Как обсуждалось выше, любое материальное тело, это динамическая система, движение элементов которой находиться за порогом восприятия наших органов чувств. Процесс развития нервной системы презентует фазу запоминания, а мышление и поведение, по своей сути, являются фазой воспроизведения.
Для обозначения качественной стороны памяти, удобно использовать представление об узоре информации, сочетании уровня возбуждения отдельных нейронов нервного узла. Характеризуя память, как целое, мы можем считать ее эквивалентом определенного состояния сознания, складывающегося из отдельных узоров информации в узлах всей нервной системы. Например, если УИ 1 рассматривать, как сочетание активности отдельных нейронов НУ 1, то дальнейшее прохождение узора информации, по цепочке дочерних НУ 2 и НУ 3, порождает в них соответственно УИ 1.2 и УИ 1.3. Оптимальной формой существования узора информации в нервной системе, являются кольцевые потоки импульсов, образуемые при участии вставочных узлов, которые направляют узор информации в обратном направлении. Градация узоров информации (в дочерних узлах), выглядит следующим образом:
УИ 1 -> УИ 1.1-> УИ 1.2 … УИ 1.Х
Соответственно, текущее состояние сознания можно обозначить, как сумму узоров информации узлов всей нервной системы, которое в свою очередь будет определятся структурами филогенетической памяти:
(УИ 1 + УИ 1.1 … УИ 1.Х) + (УИ 2 + УИ 2.1 … УИ 2.Х)
+(УИ Х + УИ Х.1 … УИ Х.Х)
Целый ряд факторов может оказать влияние на содержание филогенетической памяти. К ним следует отнести нарушения развития нервной системы в онтогенезе и повреждение нервной системы, вследствие прижизненных заболеваний. К заметным морфологическим и функциональным изменениям участков нервной системы, связанных с исследуемым сенсорным анализатором, приводит, на ранних стадиях развития организма, сенсорная депривация (64).
В плане излагаемого материала показательны опыты П.К.Анохина, по перекрестному сшиванию нервов, в результате которого два нервных узла как бы меняются местами (7). С нашей точки зрения, в результате этого эксперимента в зрелых узлах блуждающего и кожного нервов был получен процесс прижизненной перестройки филогенетической памяти, что доказывает ее пластичность, в определенных условиях. В данной ситуации можно говорить о феномене заполнения, когда общая динамическая структура нервных импульсов, помещенная в “жесткие рамки” внешней среды (наружной и внутренней), вызывает перестройку структуры нервных узлов по законам равновесия, необходимых для данного участка пространства.
15. Память мембраны нейронов
Развитие окончаний нейронов под воздействием внешней среды и заполнение ими свободных мест на суммарной мембране нервного узла, является наиболее молодой формой формообразования в нервной системе, заканчивающейся по мере роста и взросления организма. Особенности формирования нервной системы в онтогенезе можно условно выделить как особый вид памяти, память онтогенеза. В тоже время, этот вид памяти во многом совпадает с филогенетической памятью, который правильней рассматривать, как ее разновидность, определяемую прижизненными нюансами изменений среды вокруг организма.
Когда анатомические образования в нервной системе окончательно сформировались, основным фактором пластичности нейронной сети становится мембрана нейрона. При изучении нервной системы Aplisia Californica Э. Кэндел подробно описал феномен габитуации (70). Габитуация – это ухудшение ответа нейронов при однообразных воздействиях внешней среды. Кэндел различает кратковременную и долговременную габитуацию. По Э. Кэнделу, кратковременная габитуация связана с функциональной астенией пресинаптической мембраны, а долговременная обусловлена структурной перестройкой постсинаптической мембраны. В плане излагаемого материала, следует считать вполне логичным изменение структуры мембраны, под воздействием кратных внешних воздействий. Изменение микроструктуры мембраны нейрона, которое является третьим барьером противодействия внешней среде и уравновешивает избыток энергии, поступающий в нейропиль нервного узла, мы будем называть памятью мембраны.
Важно подчеркнуть, что мембрана нейрона не перекодирует энергию внешней среды, а уравновешивает ее, изменяя свою структуру! Многочисленные окончания коротких отростков нейронов диффузно располагаются по всему нервному узлу, заполняя все свободные места на поверхности его нейронов, что стирает границы между отдельными нейронами и позволяет говорить о суммарной мембране нейронов данного узла.
Процесс формирования узора информации мембраны представляется следующим образом. Энергия внешней среды, в виде узора информации, поступает в нервный узел, а оттуда в волокна нейропиля различного диаметра. Скорость движения импульса зависит от диаметра волокна и ближе к поверхности мембраны нейронов замедляется. Кроме того, особенности прохождения импульса в нейропиле определяются многочисленными эфаптическими контактами между отдельными, безмиелиновыми волокнами. В конечном счете, на суммарной мембране нейронов формируется многочисленные точечные состояния мембраны, характеризующие оттенки узора информации поступившего в данный нервный узел.
Относительно стабильный узор информации, существующий в нервном узле, получает множество оттенков, при его проецировании на его суммарную мембрану нейронов этого узла, что позволяет говорить о существовании ряда родственных узоров информации мембраны, или о памяти мембраны
В ответ на изменения внешней среды, в нервном узле возникает узор информации, характеризующий определенные условия вокруг организма. Благодаря дополнительному уравновешиванию избытка энергии, отраженного суммарной мембраной нервного узла, емкость информации, которую способен уравновесить нервный узел, значительно возрастает. Таким образом, динамическая модель внешнего мира в нейронной сети, становиться более точной. Кроме того, память мембраны, в отличие от филогенетической памяти, обладает значительно большей пластичностью и способна изменяться при жизни индивида. В местах контакта микроскопических окончаний с мембраной нейрона, величина поступающего избытка энергии соизмерима с энергией внутренних структур клетки, поэтому можно предположить, что подключение к процессу противодействия избытку энергии, поступающему из внешней среды, помимо мембраны еще и внутриклеточных структур способно сформировать внутриклеточный барьер противодействия (вероятно, этим можно объяснить феномен эйдетической памяти).

Воздействие внешней среды порождает в нервном узле узор информации, который детализируется на суммарной мембране нейронов данного нервного узла, как узор информации мембраны.
Становится очевидным, что наличие участков нейронной сети, где хорошо развита память мембраны, делает содержание сознания более стойким к повреждению, поскольку емкость узора информации значительно возрастает. В общей массе узора информации всей нервной системы, удельный вес узора информации мембраны нервного узла невелик, поэтому разрушение одного узла не может существенно нарушить содержание сознания. В развитой нервной системе, феномен заполнения более разнообразен, чем в нервной системе, в которой слабо развит нейропиль. Именно он является основным механизмом процессов ассоциирования в процессе мышления. Феномен заполнения, применительно к памяти мембраны, наглядно объясняет, почему больной, потерявший правую руку, способен писать слова на бумаге зажатым в зубах карандашом, без предварительного обучения (45).
Памяти мембраны, которая сдержится в цепочке нервных узлов, можно записать следующим образом:

Воздействие внешней среды порождает в нервном узле узор информации, который детализируется на суммарной мембране нейронов данного нервного узла, как узор информации мембраны.
Становится очевидным, что наличие участков нейронной сети, где хорошо развита память мембраны, делает содержание сознания более стойким к повреждению, поскольку емкость узора информации значительно возрастает. В общей массе узора информации всей нервной системы, удельный вес узора информации мембраны нервного узла невелик, поэтому разрушение одного узла не может существенно нарушить содержание сознания. В развитой нервной системе, феномен заполнения более разнообразен, чем в нервной системе, в которой слабо развит нейропиль. Именно он является основным механизмом процессов ассоциирования в процессе мышления. Феномен заполнения, применительно к памяти мембраны, наглядно объясняет, почему больной, потерявший правую руку, способен писать слова на бумаге зажатым в зубах карандашом, без предварительного обучения (45).
Памяти мембраны, которая сдержится в цепочке нервных узлов, можно записать следующим образом:
(УИ 1 + УИМ 1) -> (УИ 1.1 + УИМ 1.1) -> (УИ 1.2 + УИМ 1.2)
… (УИ 1.Х + УИМ 1.Х)
Соответственно содержание сознания, рассматриваемое на уровне памяти мембраны, можно записать как:
(УИ 1 + УИМ 1) -> (УИ 1.1 + УИМ 1.1) … (УИ 1.Х + УИМ 1.Х) +
(УИ2 + УИМ 2) -> (УИ 2.1 + УИМ 2.1) … (УИ 2.Х + УИМ 2.Х) +
(УИ Х + УИМ Х) -> (УИ Х.1 + УИМ Х.1) … (УИ Х.Х + УИМ Х.Х)
16. Различные подходы к изучению феномена памяти
Источником наших представлений о памяти является содержимое нашего сознания. Сознание, это динамическая структура, уравновешивающая конкретные состояния внешней среды и тканей организма. Изменение внешних условий нарушает это равновесие. Появляется избыток энергии, который нейтрализуется перестройкой ряда участков на суммарной мембране нейронов нервной системы (фаза запоминания). Повторение нестандартной ситуации встречает противодействие со стороны изменившихся участков суммарной мембраны нейронов, что определяет соответствующе содержание сознания (фаза воспроизведения). Следовательно, нервная система это хранилище различных состояний сознания, находящихся в потенциальном состоянии и способных модулировать его взаимоотношения с внешней средой, согласуясь с ранее приобретенным опытом.
К проблеме памяти обращалось большинство философских систем прошлого. В ее изучении, можно выделить два основных подхода. Первый из них, самоанализ, связан с познанием человеком самого себя. Он стал основным в философии Древней Греции и получил системное толкование в трудах Аристотеля и Платона. Самоанализ стоит у истоков возникновения большинства понятий психологии, дальнейшая систематизация которых способствовала выделению психологии из философии, в качестве самостоятельной науки. Второй метод, метод внешнего наблюдения, стал доминировать в эпоху возрождения, при переходе от созерцания к активному изучению природы. Для изучения психического мира стали использовать данные, полученные при анализе общественного поведения человека. Метод внешнего наблюдения, в том или ином объеме, использовали в своих трудах английские философы Т. Гоббс, Дж. Локк и многие другие представители их эпохи (28). Классическое обоснование метод внешнего наблюдения получил в бихевиоризме, который был разработан Э. Торндайком и Дж. Уотсоном в 19-м веке (3). С внедрением инструментальных методов исследования произошло окончательное становление психологии, как науки основанной на методе внешнего наблюдения. Например, проблемы памяти на современном этапе изучаются в клинической и экспериментальной психофизиологии, физиологии поведения, морфологии, гистохимии, электрофизиологии мозга и отдельных нейронов, аналитической биохимии и многих других экспериментальных науках.
Одним из первых философов, систематизировавших понятия психологии, был Аристотель. В трактате “О памяти и воспоминании” им представлен процесс запоминания, как появление в мозге “отпечатка” окружающего мира (23). Такой подход к рассмотрению явлений психического содержит в себе предпосылки для его изучения методом внешнего наблюдения. В тоже время Аристотель считал, у человека существует качественно новый вид памяти, называемый им воспоминанием, который связан с наличием в психике сложной системы прошлого опыта, имеющего нематериальную основу, постигаемую в процессе самоанализа.
Основным методом изучения психического у Платона, является самоанализ. В своих исследованиях он показал, что нельзя психическую жизнь человека объяснить только приобретенным опытом, поскольку значительную ее часть составляют врожденные свойства, определяемые им как “душа”. В отличие от Аристотеля, Платон рассматривал психическое, следствием активности некого идеального источника (110). В нашем понимании, врожденный опыт совпадает с представлением о той части потенциальной энергии мозга, которая сложилась в процессе эволюции нервной системы человека (с нервной тканью, как таковой). Платон не смог представить формирование “души” внешним воздействием, что послужило бы разумным компромиссом, между его взглядами и позицией Аристотеля. В тоже время, необходимо отметить, что Аристотель, и в еще большей мере Платон, рассматривали память как динамический процесс, направленный на взаимодействие с окружающим миром.
Попытку рассмотрения проблемы памяти мы встречаем в учении английского эмпиризма, которое появилось в эпоху возрождения. Основатели эмпиризма, Г. Гоббс и Дж. Локк, рассматривали логику формирования психического, в русле идей ассоцианизма (29). Сознание человека представлялась ими как чистая доска, на которую, в течение жизни, записывалось множество единичных событий. Повторное соприкосновение с этими событиями, в их неполном, редуцированном виде, вызывало воспоминание. Согласно принципу ассоциирования, возможность получить данное воспоминание, в ответ на незначительную часть ситуации в которой оно возникло, а также нарушение порядка воспроизведения воспоминаний, вызываемое изменившимися внешними условиями, определяют поступательную эволюцию психических процессов. Если к этим представлениям добавить активность самого субъекта, станет возможным объяснить такие явления, как механизмы мышления, направленность психических процессов и формирование содержимого памяти. В нашем материале, процесс ассоциирования объясняется, как следствие феномена заполнения, что глубже отражает его сущность (любое текущее воздействие внешней среды неповторимо, поэтому сознание, уравновешивая это воздействие, реагирует собственно не на него, а на целый класс, близких по структуре, внешних явлений, что воспринимается, как возникновение ассоциаций).
Ассоцианизм основное внимание акцентировал на опыте, приобретенном в процессе обучения, игнорируя существование врожденного компонента содержимого сознания. Этот недостаток, несколько односторонне, был исправлен А. Бергсоном, в работе “Материя и память”. Совершенно верно подметив, что приобретенный опыт модулируется “памятью неповторимых событий прошлого”, он определяет эти события как “образную сферу чистых воспоминаний” и “память духа” (4). А. Бергсон, в своих рассуждениях, использует исключительно метод самоанализа. Это приводит его к одностороннему выводу, что мозг является орудием проведения воспоминаний в сознание, неспособным эти воспоминания порождать или хранить (очевидный результат нежелания смотреть на психические явления глазами внешнего наблюдателя).
Несколько иное решение проблемы памяти находим в трудах З. Фрейда, создателя метода самоанализа (128). В теоретических воззрениях он полагал, что энергия подсознательного имеет образную, неизменную форму, основанную на прошлом социальном опыте (является исторической памятью событий прошлого). Поскольку структура “Я” неспособна изменяться при жизни индивида, между ней и реальной обстановкой периодически возникают несоответствия, приводящие к патологии психики. Последователи З. Фрейда предложили более пластичное понимание сущности “Я”. К.Г. Юнг характеризовал либидо как психическую энергию, отражающую интенсивность жизни индивида в различные периоды его развития. Как видим, “врожденным” оказывался лишь факт избытка энергии в нервной системе, а его структурное оформление (память), определяется текущим опытом. С нашей точки зрения, энергия подсознательного не может существовать в нервной системе, как бесформенное образование. Структура подсознательного, содержит стереотип прошлого поведения, который не вытесняется из более нового поведенческого континуума, а уточняется и совершенствуется, согласно текущим условиям существования.
В плане рассмотрения нервной системы, как фрагмента всеобщего движения, необычайно интересны идеи гештальтпсихологии, появившиеся в результате изучения особенностей восприятия движущихся объектов (4). Один из основателей гештальтпсихологии, Э.Мах, изучая психологию восприятия, обнаружил, что три нарисованные на бумаге окружности различного диаметра, которые по логике вещей должны были восприниматься как различные объекты, в нашем сознании ощущаются разновидностью единого образа “круга”. Это позволило ему ввести понятие гештальта, характеризующего наиболее простые неделимые элементы психического мира. Показательны исследования М. Вертхеймера показавшего, что восприятие того или иного объекта внешней среды зависит от характера его передвижения в пространстве.
“Если световой луч пропускался сначала через одну прорезь, а потом через другую, то при относительно длительном интервале времени (более 200 миллисекунд), наблюдатели видели последовательное появление света сначала в одной, а потом в другой прорези. Если временной интервал сокращался, то наблюдателям казалось, что обе прорези освещены постоянно. При длительности интервала более 60 миллисекунд, создавалось впечатление, что линия света непрерывно перемещается от одной прорези к другой и обратно”
Таким образом, суть гештальтпсихологии сводится к представлению о том, что движение внешнего мира способно порождать в сознании гештальты, своеобразные, неделимые кванты психического мира, которые организованы в единую систему. С нашей точки зрения, важнейшим достижением данного направления, является догадка о том, что образы сознания, порождаемые в процессе мышления или созерцания, есть реальные образования, гештальты.
Феномен формирования образа круга, при созерцании трех окружностей различного диаметра (на который обратил внимание Э. Мах), объясняется следующим образом. В первую очередь следует обратить внимание на то, что процесс зрительного восприятия активен и определяется особенностями движения глаз. Чтобы увидеть окружность, глазу необходимо научиться двигаться определенным образом. Например, ребенок в процессе своего развития вначале учится видеть круг, а потом начинает различать его разновидности (104). Слушая музыку, мы проговариваем про себя ее мелодию, усваивая ее логику (формируем моторный образ) и только потом начинаем различать ее тональные варианты. Таким образом, образ круга, восприятие мелодии, обусловлены определенным стереотипом движения глаз и голосовых связок, что и формирует в нервной системе моторный гештальт (процедурная память). Во-вторых, имеет значение сам процесс формирования образов в нервной системе. В наше сознание, при контакте с внешней средой, проникают не сами объекты, а волна избытка энергии, отражающая их динамические особенности. Циклические формы внешнего движения, индуцируют циклическое движение импульсов в нейронной сети. Если две системы уравновешивают друг друга, то кольцевые потоки импульсов, имеющие общую логику движения с внешними объектами, становятся стабильными во времени и пространстве, приобретают “образность”. Кроме того, волна энергии, продвигаясь по слоям нервной системы, испытывает на себе их фильтрующее влияние, поэтому образы сознания, по мере усложнения нервной системы, становятся все более упрощенными (абстрактными), что является “материальной базой” для появления сенсорных гештальтов (декларативная память).
По нашему мнению, гештальт следует понимать, как эквивалент кольцевого движения импульсов в нервной системе, с той разницей, что за субъективным образом не определяется само движение. Многократное прохождение импульсов по замкнутым цепям нейронов (кольца реверберации), по мнению Хебба составляющее основной механизм консолидации памяти (141), является одновременно основой существования образов сознания. Таким образом:
Гештальт – это неделимый элемент психического мира, кольцевое движение импульсов в нервной системе, потенциально представленное в сенсорном мозге, образами сознания, а в моторном, элементами поведения
Гештальтпсихология правильно наметила путь к пониманию того, что субъективная форма объектов зависит от особенностей движения составляющих их частей, но не сумела вынести эти представления за рамки психологии. Использованные ею экспериментальные методы увели проблему восприятия движения из общефилософской в узкоспециальную плоскость. Потерялись выдвинутые В. Келером идеи, что физический мир, как и мир психический, подчинен принципу гештальта. Например, в книге “Физические структуры в покое и стационарном состоянии”, Келер пишет, что процессы в коре головного мозга сходны с процессами в силовом поле. В ответ на сенсорные импульсы, в мозге возникает поле нервной деятельности, которое, подобно электромагнитному силовому полю вокруг магнита, имеет электромеханическую природу (в нашем представлении, на поверхности суммарной мембраны нейронов, существует цельная, однородная система, циклически организованных потоков импульсов, которые постоянно модулируются, как внешними воздействиями, так и свойствами самой мембраны).
Дальнейшее развитие гештальтпсихология получила в работах Курта Левина, использовавшего концепцию силового поля для объяснения поведения личности в терминах влияния на него поля общественного воздействия (теория психологического поля). Левин высказал предположение о существовании состояния баланса или равновесия между индивидуумом и его психологическим окружением. Когда это равновесие нарушается, возникает напряженность отношений, которая вызывает определенные изменения, ведущие к восстановлению баланса. Теория физических полей привела К. Левина к мысли, что психическая деятельность человека происходит в условиях воздействия психологического поля, которое получило название годологического пространства, формируемого личностными потребностями человека во взаимодействии с его психологическим окружением. Как видим, в логически верной концепции Левина, нет окончательного вывода о том, что феномен поля сознания, это определенный уровень организации материи, в котором существуют свои законы движения, отличные от законов существовавших на предыдущих уровнях ее организации.
Концепцию, напоминающую теорию психологического поля выдвинул немецкий психолог Э. Геринг и его последователи (Р. Земон, Э. Блейер). Созданное ими учение о “мнеме” рассматривали память не только как психическую, но и как “общеорганическую функцию”. К сожалению, стереотипное мышление не позволило этой гипотезе перерасти в более широкую систему взглядов.
Французская школа социологии (П. Жанне) обратила внимание на историческую природу и социальную обусловленность памяти человека. Согласно ее представлениям, память это процесс, действие “специально изобретенное людьми”, принципиально отличное от простой репродукции (126). Как видим, данная концепция акцентирует внимание на активной стороне памяти, фазе воспроизведения. Если развить эти взгляды далее, то память надо рассматривать в парадигме категории взаимодействие, а “коллективную память”, как проявление общей, динамической картины мира, которая модулируется участием элементов социума. Похожие взгляды встречаем у английского психолога Ф. Бартлетта, по мнению которого память является схематической реконструкцией прошлого опыта, где важна не точность воспроизведения информации, а ее глубинный смысл или общая направленность всего процесса (127). Подобное понимание памяти подразумевает рассмотрение ее в качестве проявления реально существующей динамической системы, а не склада информации. Однако оба подхода, предложенных к решению проблемы памяти, должны были бы опираться на соответствующую философскую систему, которая легализовала бы их существование.
В советской школе психологии теоретические вопросы изучались с точки зрения диалектического материализма. Так в работах Л.С. Выготского, А.Н. Леонтьева, П.И. Зинченко, А.А. Смирнова, память рассматривалась, как динамическая структура, процесс, связанный с предметной деятельностью человека. В целом такой подход не встречает возражений, но требует детализации применительно к экспериментальным данным.
Клинические исследования Т. Рибо доказали, наличие таких разновидностей памяти, как память-привычка (моторная память), эмоциональная или аффективная память (память чувств) и словесно логическая память, нарушение которых совпадало с локальными повреждениями определенных зон мозга (10). Отсюда следует вывод, что память имеет локализацию и определяется не только микроструктурами мозга, но и его макроструктурами. По нашему мнению, компромисс между сторонниками локального и диффузного представительства памяти решается следующим образом. Память (энграмма) доступна нам частично, как содержание нашего сознания, временно стабильное состояние кольцевых потоков импульсов в нервной системе. В формировании сознания, так или иначе, участвуют все образования мозга. Информация, поступившая в сознание, как результат раздражения определенного участка мозга, становиться частью общей динамической системы и теряет свою самостоятельность. Вопрос о локальных структурах, оказывающих влияние на процесс запоминания или воспроизведения, должен рассматриваться в контексте дефекта, который наноситься общему объему энграммы, всему полю сознания. Любое повреждение нейронной сети компенсируется динамикой сознания, как цельной системы и зависит не только от состояния мембраны нейронов, но и от условий внешней среды. Таким образом, участие тех или иных структур в формировании памяти, необходимо рассматривать в более сложной, чем это принято, системе координат.
Современные подходы к изучению памяти опираются в основном на данные, полученные в эксперименте, с использованием разнообразной инструментальной базы. Многие положения о сущности памяти и механизмах ее формирования, существующие в современной психофизиологии, близки к тем, которые излагаются в данной работе. В тоже время, обширность и разнообразие экспериментального материала затрудняет широкие обобщения. Например, Т.Н.Греченко пишет (12):
“Развитие концепции временной организации памяти затрудняется нечеткостью и расплывчатостью основных понятий, на которые опирается теоретическая конструкция”
Используя наш подход к рассмотрению проблемы памяти, можно яснее представить некоторые теоретические вопросы психофизиологии. Широко распространена точка зрения, согласно которой, временная организация следа памяти подразумевает последовательность развития во времени качественно разных процессов, приводящих к фиксации определенного опыта. На основании изучения последствий применения различных амнестических агентов, выделяют память долговременную и кратковременную. Такой, дискретный подход, к пониманию памяти, уводит в сторону от ее сути, как процесса, характеризующего взаимодействие мозга с окружающей средой.
О содержании памяти мы судим по состоянию посредника, поля сознания, которое определяется структурой нейронной сети и особенностями внешнего окружения. Особенности нейронной сети, которые в свою очередь обусловлены прошлым опытом взаимоотношений сознания с внешней средой, определяют потенциальную возможность памяти влиять на поле сознания. Макроструктура нервной системы создает грубую копию окружающего мира, благодаря которой мы обладаем способностью недифференцированно воспринимать различные параметры внешней среды: видеть, слышать, осязать. Макроструктура нервной системы, которая развилась в процессе ее эволюции, не способна фиксировать прижизненный опыт. Запоминание текущих событий и оттенков образного восприятия, становятся возможными благодаря появлению прижизненной пластичности микроструктуры мембраны нейронов.
Потоки импульсов на мембране нейронов, не есть структура памяти, они представляют “передний край” внешней среды (являются посредником между внешней средой и мембраной нейронов). Применение электрошока нарушает кольцевые потоки импульсов в нейронной сети и не затрагивает “оправу”, в которую они заключены (способность воспринимать образы внешнего мира нарушается только при органических дефектах нейронной сети). Разрушение поля сознания прерывает процесс фиксации его содержания на суммарной мембране нейронов (место электрошока, с тем же успехом, можно было бы поставить экран на пути зрительного анализатора, уменьшив экспозицию внешних воздействий на мембрану нейронов до такого уровня, когда зрительный образ неспособен запечатлеться). Поскольку процесс консолидации растянут во времени и зависит от стабильного существования поля сознания, он по-разному чувствителен к амнестическим агентам в зависимости от их качества, времени использования, места применения и так далее. В результате создается впечатление многоплановости процесса запоминания. Таким образом:
Кратковременная память, это условие для формирования памяти в нейронной сети, временно стабильное состояние поля сознания, при котором мембрана нейронов еще не успела выработать механизмы, уравновешивающие данное воздействие внешней среды
Долговременная память проявляется в потенциальной возможности нейронной сети влиять на содержимое поля сознания, а ее субстрат соответствует всей массе мозга
Гипотеза о двух последовательно развивающихся следах памяти, предложенная Д.О. Хеббом и Р.В. Джерардом, ошибочно подменяет условия для процесса запоминания, понятием “кратковременная память” (47). В данном случае, речь может идти не о механизмах кратковременной памяти, а о том, что наличие колец реверберации характеризуют память поколений, результат эволюционного развития вещества мозга. Наличие колец реверберации создает среду, через которую становиться возможным запоминание прижизненных событий.
Гипотеза одного следа и двух процессов Дж. Л. Мак-Го и П.Е. Гоулда предполагает, что процесс формирования кратковременной памяти находиться в зависимости от общего состояния нервной системы, так называемых неспецифических явлений, изменения уровня бодрствования, уровня определенных гормонов и так далее. Как видим, линия рассуждений далека от представления, что процесс развития мозга, одновременно является процессом формирования субстрата памяти. Противоречит данной гипотезе факт восстановления под гипнозом незначительных деталей прошлого, которые, с ее точки зрения, не должны были бы запоминаться вообще. Противоречит ей и феномен абсолютной памяти гражданина Ш., описанный у А.Р. Лурии (86). С нашей точки зрения, память – это способность ткани мозга влиять на состояние поля сознания. Структура сознания модулируется внешними процессами в среде вокруг организма и внутри его. Само собой разумеется, что “информация” от тканей организма (в том числе и воздействие медиаторов, гормонов), всегда будет участвовать, в формировании содержания сознания, по которому мы судим о проявлениях памяти. Таким образом, гипотеза одного следа и двух процессов не столько создает концепцию памяти, сколько констатирует факт участия активирующих систем мозга, в процессах ее формирования. Поскольку мотивация активирует ориентировочное поведение, в формировании энграммы участвует большое число систем мозга, а ее содержимое, даже в редуцированном виде, легче воспроизводиться.
Наша концепция проясняет ряд фактов, плохо объяснимых гипотезой временной организации памяти. Например, спонтанное восстановление старых следов памяти, после действия химического амнестического агента, можно объяснить несколькими причинами. Во-первых, амнестический агент создает в поле сознания избыток энергии, который в первую очередь повреждает мембрану нейронов, реагирующих на изменившиеся внешние условия, так как их активность близка к максимальному значению. Во-вторых, приобретенные навыки реализуются в поле сознания, структура которого, прямо или косвенно, определяется всей нервной системой, поэтому искажение свойств небольшого участка мембраны, где храниться вновь приобретенный след памяти, не может грубо нарушить общее содержание энграммы. Таким образом, избыток энергии, привнесенный экспериментатором в нейронную сеть, в условиях, когда внешние воздействия практически восстановили основную массу энграммы, заставляют поврежденный участок мембраны изменятся по логике близкой к логике ранее сформированной энграммы. Иными словами, любой избыток энергии в нервной системе активирует стандартные механизмы противодействия мембраны нейронов, находящиеся в латентном состоянии. Если эти механизмы разрушены или повреждены, а общая логика динамики процессов сознания, в соответствующих внешних условиях, сохраняет большую часть поврежденной энграммы, то поврежденные участки мембраны приобретают свойства близкие к тем, которые были утрачены ранее.
Из сказанного следует, любой избыток энергии в нервной системе распределяется по ее структурам и активирует механизмы противодействия мембраны нейронов, то есть процесс воспроизведения, который по своей сути ближе к феномену заполнения (описанному выше). Воспроизведение каждый раз происходит по-новому, что связано с неповторимостью внешних условий, повреждением мембраны нейронов и так далее. В тоже время, если механизмы воспроизведения нарушены или повреждены, то общая логика процессов сознания сохраняет определенный стереотип действий, связанный с прошлым опытом, который составляет значительную часть любой энграммы. Этот же стереотип определяет вектор движения нервных процессов и в случае повреждения участка мембраны или появлении внешних условий близких к тем которые существовали ранее, приводит к формированию в сознании энграммы, близкой к определенным стандартам (поле сознания объединяет всю нейронную сеть в единую динамическую систему и поврежденный участок мембраны восстанавливает свои свойства в контексте ранее сформированного динамического стереотипа). Таким образом, свойства любого участка мембраны нейронной сети находятся в зависимости от состояния остальных ее участков, а процесс воспроизведения соответствует стремлению взаимодействующих систем к равновесию.
Руководствуясь этой логикой, можно объяснить, как происходит восстановление следа памяти методом напоминания, после его депрессии электрошоком, когда перед тестированием животным предъявляли слабое электрическое раздражение, которое, по сути дела, создавало избыток энергии в нервной системе и тем самым формировало новую энграмму, близкую по содержанию к энграмме, существовавшей ранее. Параллельно можно объяснить механизм восстановления следа памяти при двойном электрошоке (в данном случае имеет место тот же избыток энергии, величина которого частично нивелируется формированием механизмов противодействия мембраны при первичном воздействии, в результате чего мы получаем вышеописанную ситуацию).
Нетрудно объяснить механизм восстановления энграммы в экспериментах по методу ознакомления. Если перед обучением животное поместить в экспериментальную камеру и дать возможность свободно по ней передвигаться, то, после применения амнестического агента, ретроградная амнезия не возникает. Фактически, при ознакомлении происходит значительное расширение содержания будущей энграммы и разрушение вновь приобретенного опыта составит меньшую часть, от общего ее объема, в сравнении с таковым без ознакомления.
Экспериментально установлено, если после перехода памяти в долговременное хранение производилась реактивация энграммы методом “напоминания”, то в создавшихся условиях старый след памяти становился уязвимым для электрошока. Был получен вывод, что для амнестического воздействия досягаемы следы памяти, при его применении находящиеся в активном состоянии. Согласно нашей точке зрения, при использовании, электрошока “энергетический удар” приходится по тем нейронам, которые “уравновешивают” данное содержание сознания и уровень возбуждения которых близок к максимальным значениям. Поступивший в мозг избыток энергии заставляет нейроны ему противодействовать и повышать свой уровень активности, при этом нейроны, изначально обладавшие более высоким уровнем возбуждения, будут больше уязвимыми к энергетическим воздействиям. Ретроградная амнезия, которая развивается после механической травмы мозга или электрошока, обусловлена, с одной стороны, разрушением сознания и искажением информации, поступающей на активные участки мембраны, а с другой, связана с изменением свойств самой мембраны, в результате локального появления избытка энергии.
Концепция активной памяти, совершенно справедливо рассматривает ее как нечто целое, без разделения на память кратковременную и долговременную. В тоже время, с нашей точки зрения, утверждение, что память существует в активной форме, готовой для реализации в данный момент времени и в пассивной форме, не готовой к непосредственному воспроизведению, нуждается в переосмыслении (120). Память не может быть активной или пассивной. Иллюзию активности создает перераспределение силы пульсации отдельных нейронов, при поступлении избытка энергии в поле сознания (часть суммарной мембраны нейронов изменяется, уравновешивая дежурное состояние сознания). Если основная масса нейронной сети сохраняет свое исходное состояние, являясь потенциальной частью памяти, это не значит, что пассивная память не участвует в формировании поля сознания. “Пассивные” участки мембраны создают условия, без которых невозможно существование поля сознания в состоянии, способном воспринять текущую информацию. Нельзя говорить, что пассивная часть памяти готова к реализации, она постоянно находиться в этом состоянии, создавая динамический фон, на котором возможно участие специализированных участков мембраны в формировании поля сознания.
Гипотеза о распределенности энграммы утверждает, что есть структуры мозга привязанные к кратковременной и долговременной памяти (136). По нашему мнению, данное предположение основано на непонимании, что кратковременная память это собственно не память, а состояние сознания, создающее условия для процесса консолидации, характеризуемого как долговременная память. Нередко формируется мнение о существовании ограниченного объема кратковременной памяти, хотя речь должна идти о том, что ограниченным является не объем памяти, а количество информации, способной поступить по сенсорным каналам в поле сознания.
Непосредственное отношение к памяти имеют проблемы научения, которые изначально рассматривались, как предмет педагогики. Позднее, исследование научения у животных, с использованием нейрофизиологических методик, способствовало появлению отдельного направления психофизиологии, занимающегося этими вопросами. В парадигме философии движения, поведение, это вектор движения организма во внешней среде, направленный на достижение с ней равновесия. Механизм поведения реализуется автоматически, как следствие объединения в поле сознания сенсорной информации от внешней среды с прежним опытом активности тканей организма, представленной обобщающим образом нашего “Я” (“темное мышечное чувство” по И.М. Сеченову). Фактором эволюции нервной системы и поведения, является избыток энергии, обусловленный периодически возникающим нарушением равновесия с внешней средой (появившийся в мозге избыток энергии порождает динамику состояний сознания, что обеспечивает выработку новых программ поведения и адаптацию организма к изменившимся условиям внешней среды).
Близка к нашим преставлениям идея К. Прибрама о том, что некоторые участки нервной системы напоминают голографическую пластину (106). Однако, с нашей точки зрения, такой пластиной является вся ткань мозга, а не только нейропиль, а динамика состояний сознания, под воздействием энергии сенсорного воздействия, соответствует процессу воспроизведения “голографического образа”. Экспериментально доказанное “плавание энграммы по структурам мозга” вполне логично, если учитывать постоянно меняющиеся взаимоотношения организма с окружающей средой и стремление взаимодействующих систем к равновесию. Можно разъяснить парадокс, с которым столкнулся К.С. Лешли, утверждавший, что памяти нигде нет, но в тоже время она всюду (93). Память, это дежурное состояние сознания, в формировании которого принимает участие вся нервная система. Полное разрушение сознания соответствует повреждению всего мозга, как целого. Локальное нарушение любого участка нейронной сети изменяет качество “дежурной” энграммы (значимость дефекта, при повреждении ткани мозга зависит от его участия в данном, конкретном содержании сознания). При участии в создании энграммы участка нервной системы, подвергнутого локальной деструкции, в поле сознания поступает волна изменений, которая перестает принадлежать данному участку мембраны и становиться частью целого, разрушить которое оно не в состоянии.
Деление памяти на процедурную и декларативную роазновидности, относительно. Еще большим заблуждением является представление о принципиальном различии этих видов памяти. В нервной системе просматриваются два функционально отличных отдела, сенсорный и моторный мозг. Преставление о процедурной памяти (знание того, как нужно действовать), связано с активностью моторного мозга, образования которого уравновешивают мышечную и гормональную системы организма. Особенности декларативной памяти (которая обеспечивает ясный и доступный отчет о прошлом индивидуальном опыте) обусловлены сенсорным мозгом, структуры которого уравновешивают потоки импульсов, поступающие через сенсорные анализаторы из окружающей среды. Поскольку содержимое декларативной и процедурной памяти, поступая в поле сознания, теряет свою самостоятельность, становясь частью общей динамической системы, их разделение относительно.
Большинство работ по психофизиологии нервной системы, касаются отдельных ее структур. В то же время существуют подходы, позволяющие рассматривать мозг, как единое целое, где нервная система, вольно или невольно, рассматривается в виде неорганической системы, к которой применимы физические законы. Например, Е.Н. Соколов, создатель векторной психологии, показал, что движение импульсов в нервной системе можно описывать, как движение дискретных элементов в неорганическом мире, с использованием соответствующего математического аппарата(120). П.К. Анохин, в теории функциональных систем, рассматривает вопросы организации и функционирования нервной системы в парадигме системного подхода, который выходит за рамки экспериментальной нейрофизиологии (7). Определенный шаг, в этом направлении, делает “философия движения”, которая определяет место нервной системы в общей организации материи, что открывает новые возможности для интерпретации всего комплекса научных данных. Н.П. Бехтерева использовала, для описания процессов происходящих в нервной системе, представление о паттерне импульсов, называя их нейронными кодами (21). В нашей работе тоже возникла необходимость в использовании понятия, близкого, по сути, к паттерну импульсов, которое получило определение “узор информации”.
Показательны в этом плане выводы М.Н. Ливанова, который на основании анализа данных электроэнцефалографии, сумел выявить ряд универсальных закономерностей, одной из которых является константа Ливанова. В 1938 году М.Н. Ливанов писал (47):
“Достаточно возникнуть расхождению между частотой раздражения и частотой мозгового ритма на 1/10 долю периода, как прежний корковый процесс замещается новым”
По нашему мнению, универсальность константы определяется тем, что нейронная сеть и поле сознания образуют сравнительно однородную систему, внутри которой формируются уровни движения. Как показано в данной работе, формирование нервной системы происходит в рамках усложнения циклически организованной динамической системы. Поступление избытка энергии в такую систему компенсируется, до определенного уровня, нарастанием скорости циркуляции составляющих ее элементов. Если избыток энергии превышает определенный порог, то происходит образование новых уровней движения. Вероятно константа Ливанова, определяемая как 10% новизны, ведущей к изменению характера электроэнцефалограммы, соответствует величине избытка энергии способного нарушить равновесие циркуляции импульсов в нервной системе и привести к перестройке существующего, или образованию нового, уровня движения. Многоуровневую организацию функциональных процессов в нервной системе предвидел Н.А. Бернштейн, который утверждал, что существуют неокинетические процессы (нейронная импульсация), появившаяся в филогенезе довольно поздно и палеокинетические (медленные колебания мембранных потенциалов), по происхождению более древние, метаболические, свойственные не только нейронам (18).
Известный этолог Л.В. Крушинский выдвинул гипотезу об уровне возбудимости мозга, как модификаторе поведения животных. Эта гипотеза основывалась на результатах опытов, где искусственное повышение возбудимости нервной системы введением фармакологических средств у трусливых и злобных собак, приводило к значительному усилению активных и пассивных оборонительных реакций (67). По нашим представлениям, возбудимость мозга должна рассматриваться в контексте избытка энергии, поступающего в нервную систему при нарушении ее равновесия с внешней средой. Как видим, множество отдельных гипотез и догадок, о процессах протекающих в мозге, обесценивается рассмотрением их в узкоспециальных рамках.